
Introduction (1.0.0)
Vertebrata is traditionally a subphylum. Given the arrangement in this text not all vertebrates have vertebrae. However, there is another arrangement of vertebrates which moves one group described here out of the Vertebrata and creates a clade Craniata to contain both this group and the vertebrates. This is described more in the section on the Cyclostomata.
The first vertebrate was probably a suspension-feeding, vaguely-fishlike organism that swam. The exact timing of the separation between the vertebrates and the tunicates remains unclear, with estimates ranging between 800-550 million years ago (Blair & Hedges, 2005; dos Reis et al., 2015).
This section is arranged in a traditional jawless fish to mammals arrangement, despite the fact that mammals have no automatic claim to being “most advanced”. (Indeed, even by the standards which are usually used to claim this place for mammals birds have a better claim.) This requires discussing groups that split off of the lineage that leads to mammals. While some of these groups may be termed “basal” I have opted for the less-controversial terminology of sister groups. For instance, the first group to split from the lineage that leads to the rest of the vertebrates is the Cyclostomata. This can be described by saying that the Cyclostomata are sister to all other vertebrates. This sister group naming means two things: first, all other vertebrates are more closely related to each other than they are to the cyclostomes (all other vertebrates are monophyletic) and second, the cyclostomes are the organisms most closely related to all other vertebrates.
For each group the approximate number of species is listed. The species number is drawn from Species 2000 & ITIS Catalogue of Life (Roskov et al., 2018). It is impossible to keep a completely accurate tally of all named species on earth, and so I have made the choice to draw all numbers from one source for consistency.
The traditional Linnaean ranks of many of these groups are listed as well.
Cyclostomata (1.0.0)
(Traditionally a Superorder, approximately 129 species, comprising 0.19% of all vertebrate species)
The Cyclostomata is a group of jawless fish which is sister to all other vertebrates (which are grouped together in the Gnathostomata). It includes two separate lineages of fish within it, the lampreys and the hagfish. The separation between lampreys and hagfish is very old. Molecular clock dating of the lamprey-hagfish split consistently estimates it to be at least 400 million years old, potentially an event that only briefly followed the split between cyclostomes and gnathostomes (dos Reis et al., 2015; Kuraku & Kuratani 2006).
Cyclostomes are most notable for the characteristics that they lack, which includes jaws (cyclostomes means “circle mouths” because of this), myelin (Bullock et al., 1984), and paired fins. Cyclostomes also have cartilaginous skeletons.
Controversy
The existence of Cyclostomata is debated. The cyclostome hypothesis (preferred by this author) states that hagfish and lampreys are each other’s closest relatives and that the “primitive” character of hagfish is the result of character loss in hagfish and/or independent character gain in lampreys. For instance, reduction in the eyes of hagfish seems to correlate with habitat, allowing for the possibility that all hagfish evolved from something with better vision (Fernhom & Holmberg, 1975).
The alternate hypothesis proposes that hagfish are the sister group to the other vertebrates, which are labeled as the true vertebrates as hagfish do not actually have vertebrae. The vertebrate + hagfish clade then becomes Craniata, named after the cranium found in all members. Under this hypothesis lampreys are the sister group to the gnathostomes, making them more related to Gnathostomes than to hagfish.
In general, morphological studies have tended to support the hypothesis that hagfish are sister to the vertebrates and molecular studies have tended to support the existence of Cyclostomata (Heimberg et al., 2010). This text follows the cyclostome hypothesis in large part because I believe we have reached the point where molecular phylogenies are more accurate than morphological phylogenies.
Myxini (traditionally a Class, approximately 82 species)
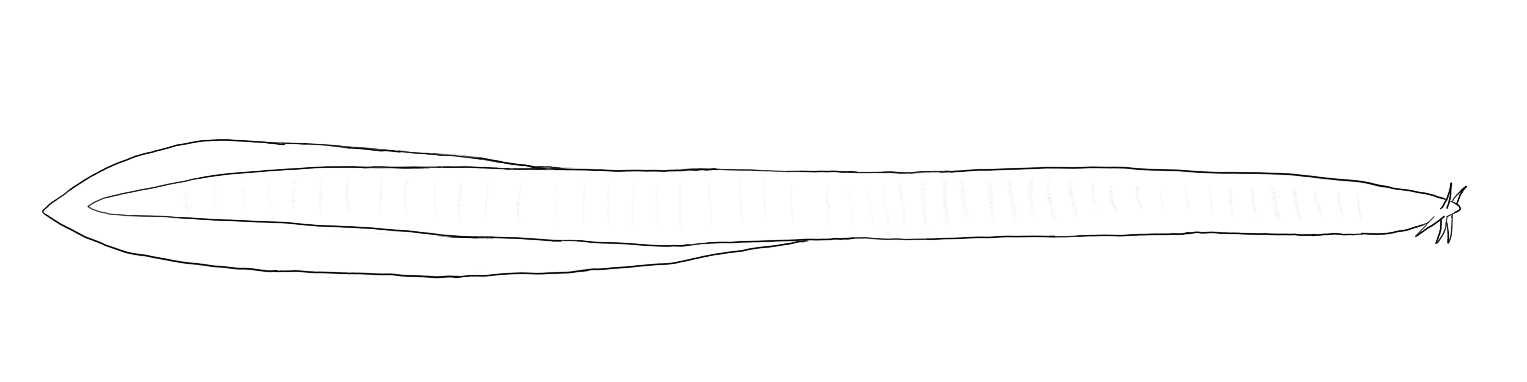
The hagfishes (Myxini) are most notable for their extremely primitive appearance. In addition to the cyclostome lack of jaws and paired fins hagfish have (depending on species) very simple eyes or no eyes at all. Unlike lampreys, hagfish have no visible gill pores and have six oral tentacles.
Bardack (1991) describes a fossil hagfish from the Carboniferous period, some 300 million years ago. This hagfish is substantially similar to modern hagfish, indicating that this basic body plan has been successful for an extremely long time.
Modern hagfish are carnivorous and, despite the absence of jaws, can bite food with movable tooth plates, producing forces that are comparable to those of many jawed fishes (Clark & Summers, 2007). Hagfish can also be observed tying their bodies into overhand knots and pulling their heads through these knots to pull at food. Hagfish are were first described as scavengers and are certainly common at baited sites along the seafloor and at whale falls. However, stomach contents show that hagfish eat marine worms, crabs, echinoderms, and shrimp, many of which are assumed to be actively preyed upon (Shelton, 1978; Zintzen et al., 2011; Zintzen et al., 2013). Zinzten et al. (2011) observed, via remote video, a slender hagfish (genus Neomyxine, species unknown) successfully hunting and eating a small burrowing fish. The slender hagfish on the video frequently investigated burrows and burrowed through the mud themselves, suggesting active hunting for buried prey. Hagfish are also known to finish off fish caught in nets, sometimes burrowing inside their bodies and eating them from the inside. However, while hagfish share dietary similarities different species appear to differ in the extent to which they rely on scavenging versus hunting (Zinzten et al., 2013).
Hagfish are also able to produce copious slime from a number of pores on the body. This slime appears to have an anti-predator function and in remote video observations numerous species of predatory fish have been observed attacking hagfish, which then produce slime, causing the predatory fish to gag and spit the hagfish out of their mouths. In some cases the predator can be seen continuing to “spit” after the hagfish has been expelled. It is likely that the slime, which is formed by proteins that bind to water, causes suffocation when inhaled into the gills (Zinzten et al., 2011).
Hagfish are generally bottom-dwelling and always marine, and in some areas can be extremely abundant.
Their reproduction is poorly studied, although they are known to be oviparous and lack larval stages.
Hyperoartia (traditionally a class, approximately 47 species)
The lampreys have been classified as Cephalaspidomorphi, Petromyzontida, and Hyperoartia in recent years, depending on how various scientists have interpreted the relationship of modern lampreys to extinct jawless fish.
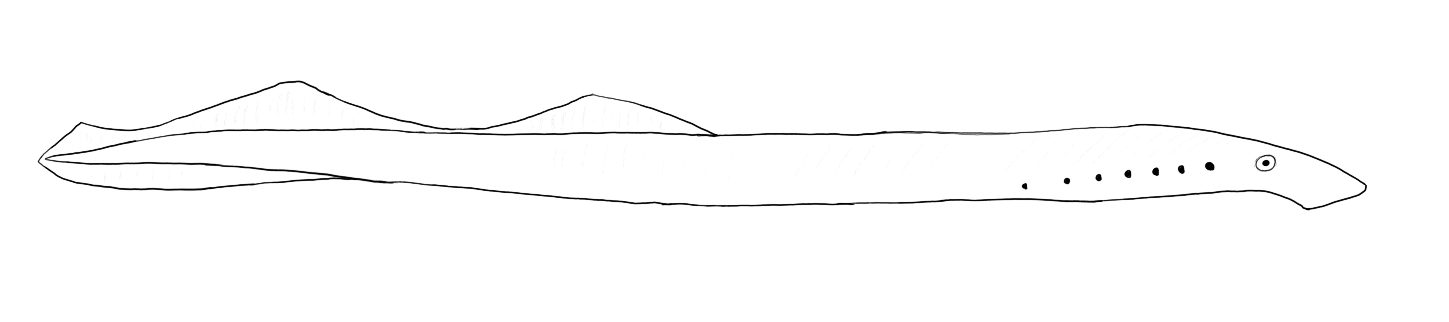
Adult lampreys looks distinctly more fish-like than hagfish, with visible eyes, gill pores, and a single nostril. Unlike hagfish, they do not bite with tooth plates but with a toothed “tongue” that slides up and down the pharynx to shred flesh (Hilliard et al., 1985).
Like hagfishes, lampreys have ancient fossil ancestors that closely resemble the modern forms. The oldest of these is the 360 million year old fossil discovered by Gess et al. (2006).
Lampreys have a more complex life cycle than hagfishes. Adult lampreys use pheromones released by larval lampreys to home in on rivers in which larval lampreys are surviving and growing (Twohey et al, 2003). Nests are built in gravel by the adults, which use their oral disk to move stones (Stone & Lohman, 2006). Eggs are laid in the nests and hatch into ammocoetes larva. These larva closely resemble cephalochordates and, like them, are suspension feeders that pump water across their pharynx and use mucus to trap fine organic matter (Mallatt, 1981). Ammocoetes larva move from the spawning sites to areas with finer sediment which they burrow into. Eventually, the larva metamorphose into adults.
Adult lampreys fall into two categories: in some species the adults do not eat and the adult portion of the lifecycle is quite short. In other species the adults are parasites and use their oral disks to attach to fish which they rasp open wounds into. In some of these species the lampreys only consume blood but in other the lampreys also consume flesh. While these lampreys are generally considered parasitic a sufficiently large lamprey can incidentally kill a fish by feeding on it. In most parasitic species the adults migrate to the ocean, but some populations have become landlocked and spend their entire lives in freshwater.
While many lamprey species are endangered the sea lamprey (Petromyzon marinus) is considered a pest in the Great Lakes. Because of this it is also intensively studied and much of what we know about lampreys comes from this species.
Gnathostomata (1.0.0)
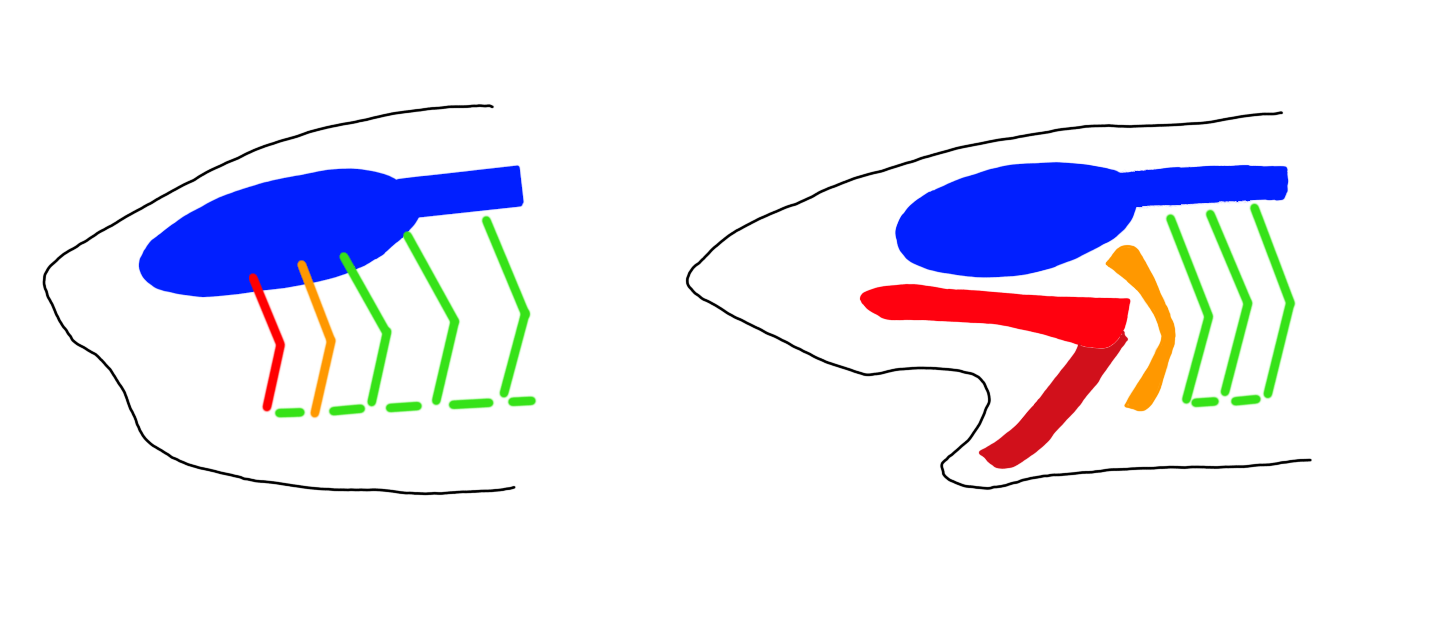
All jawed vertebrates (99.8% of all vertebrates) are gnathstomes, which means “jaw mouths”. While hagfish show that an animal can develop a powerful bite without jaws the jawless bite of a hagfish is slow (Clark & Summers, 2007). Modern vertebrate jaws allow for a faster bite, improving the efficiency with which a predator can grab a prey item. In water, where jawed vertebrates evolved, the ability to rapidly open the jaws is important for suction feeding, in which prey is sucked into the mouth by the movement of water. This method of feeding is extremely common in jawed fish.
The first step towards jaws was the evolution of the gill arch. The pharyngeal bars of the vertebrate ancestor had served a double function, both filtering food from the water and providing a respiratory surface. In vertebrates the respiratory function took over and the pharyngeal bars became the gill arches that support the gills. In the evolution of jaws the first gill arch became a jointed structure that formed the jaws. The second gill arch also lost its function as a gill arch and became involved with the musculature of the jaws as the hyoid apparatus (Kuratani, 2004).
Early jawed fish did not have fully “modern” mouths. The giant predator Dunkleosteus terrelli did not have teeth but, instead, a sharpened, spiked edge on the bones that formed the jaws. Despite this it had an impressive bite, generating more than 5,000N of force, one of the most powerful bites of any animal (Anderson & Westneat, 2007).
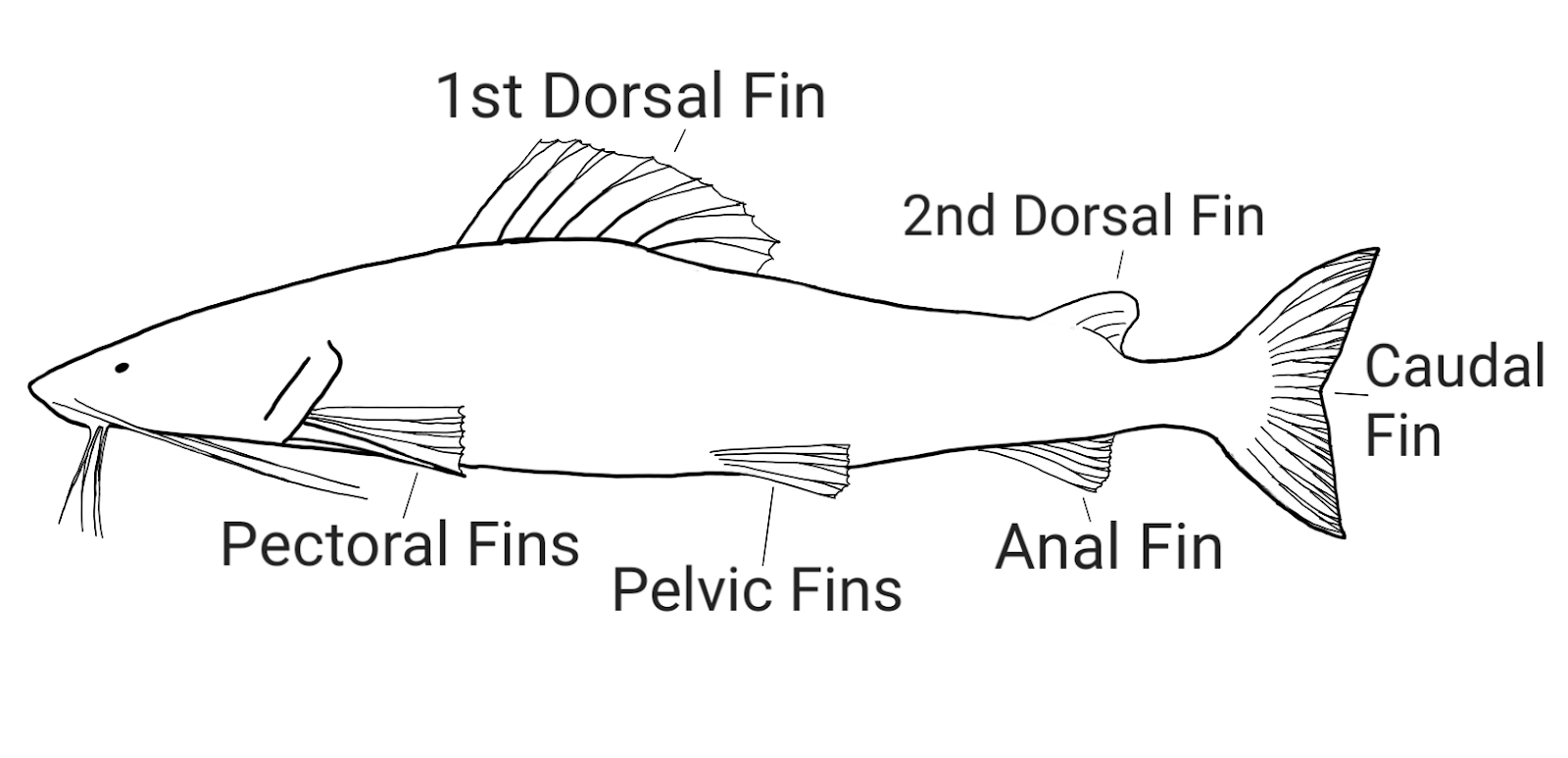
All gnathostomes also share a paired fin body plan. Cyclostomes have only unpaired fins that run along the midline of the body. Gnathostomes developed a body plan with three or four unpaired fins (one or two dorsal fins on the dorsal midline, a caudal fin on the tail, and an anal fin on the ventral midline) and two pairs of paired fins. These paired fins, the pectoral and pelvic fins, are retained as the limbs in the tetrapods. Paired fins probably allow for better stabilization in high-speed swimming.
Chondrichthyes (1.0.0)
(Traditionally a Class, approximately 1,282 species, 1.92% of vertebrate species)
The Chondrichthyes are the cartilaginous fish (which is what their name means). All living members have cartilaginous skeletons, some with interesting adaptations to generate allow skeletal elements to take higher loads despite being made of cartilage (Summers, 2000). Calcification does occur in the cartilage of the skeleton (Dean & Summers, 2006) and the lack of bone in the Chondrichthyes is almost certainly a secondary loss of bone as the most plausible ancestors of modern Chondrichthyes had bone.
Modern chondrichthyans tend to have fleshier fins than most other modern fish, with fin rays buried under tissue. The classic triangular shark dorsal fin is a good example of this, with the supporting rays completely covered.
Chondrichthyans are sometimes portrayed as “living fossils”, unchanged since ancient times. While there are ancient chondrichthyan fossils that could be superficially mistaken for modern sharks and the fossil record of the chondrichthyans includes good fossils from the early Devonian (Miller et al., 2003) there are also extremely unusual extinct chondrichthyans with no living relatives, like Heliocoprion, a shark-like animal with a whorl of teeth that resembles a circular saw. Modern sharks and rays all belong to a group called the Neoselachii which diversified rapidly during the Jurassic and Cretaceous, a time at which many modern groups of animals appeared. Unfortunately, the lack of bone in chondrichthyans means that they fossilize poorly, and so many fossil species, including the famous megalodon shark, are known entirely or almost entirely from teeth.
All known chondrichthyans are carnivores, although their carnivory includes suspension-feeding by large elasmobranchs on small fish and crustaceans, active hunting of large fish and marine mammals, and more sedentary predation on benthic organisms.
Chondrichthyans are all internally-fertilizing animals. In the males the pelvic fins are modified to form organs called claspers which are used to inseminate the female.
The modern chondrichthyes come in three major body forms: the chimeras or ratfish, the sharks, and the rays. The sharks and rays are related to one another and there are some intermediate body forms in the this group.
Holocephali
The holocephali are a small group of mostly deep-sea fish with long, thin tails, large heads (and often eyes), and an otherwise shark-like arrangement of fins. Some have a prominent dorsal fin spine which may be venomous (Halstead & Bunker, 1952). Unlike the elasmobranchs, holocephalans have gill covers and their gill openings are hard to see, being located near the pectoral fin articulation. They are known as chimeras, ratfish, rabbitfish, or elephant fish (in species with long rostrums). Because they are not a very diverse group and are a mostly deep-ocean organisms they are poorly known and many new species probably await discovery.
We do know that holocephalans are carnivores that feed on a variety of small organisms, including some fish (Giacomo & Perier, 1996). Holocephalans have “plates” of teeth that form crushing surfaces that allow them to eat hard-shelled bivalves and gastropods (Huber et al., 2008).
Modern holocephalans are oviparous (Wourms, 1977) but fossils indicate that some fossil forms gave birth to live young that fed on either unfertilized eggs or on their siblings while still in the mother’s body (Lund, 1980).
Elasmobranchii
Most living chondrichthyans are elasmobranchs which are split between two body forms. Unlike in some other groups these body forms are also clades. The sharks are generally long-bodied and strong swimmers. In sharks the gills slits are clearly visible between the head and the pectoral fins, which, like most of a shark’s fins, are triangular. The tails of sharks are heterocercal, meaning that the upper lobe of the caudal fin is longer than the lower lobe. This provides lift, and in the thresher shark the extremely elongated upper lobe of the caudal fin is used to stun small fish (Aalbers et al., 2010).

The other body form seen in elasmobranchs are the rays. Rays are dorso-ventrally flattened, with hugely enlarged pectoral fins the form a roughly disk-like shape along with the body. The pelvic fins are reduced and the dorsal fins and anal fins are absent. The caudal fin forms a slight leaf-shape around the end of the tail. The gill openings are on the ventral surface of the body, along with the mouth. Since the ray body shape is adapted to allow rays to bury themselves in sediment on the seafloor the gills are often buried and the ray respires through the large spiracles (additional openings to the gill chamber) located posterior to the eyes. While the stingrays are well-known for their venomous barb, located near the base of the tail, this is not a characteristic of all rays.
A few elasmobranchs break these rules, with species like the sawfish and sawshark appearing to be partially-flattened sharks (both with long rostrums armed with saw-like teeth). The orectolobid sharks spend much of their time on the seafloor and are also more dorso-ventrally flattened than many other sharks. Conversely, while the ray body form is adapted for living on the seafloor the manta rays are pelagic swimmers that filter food from the water column.
Elasmobranchs can grow quite large. Numerous shark species grow larger than human beings and the largest elasmobranch, the whale shark (Rhincodon typus) can easily grow to 12 meters (39 ft) in length as an adult (Graham & Roberts, 2007) and probably tops out at 18.8 meters (61.7 ft) (McClain et al., 2015). The great white shark (Carcharodon carcharias) is the largest predatory shark and is significantly smaller. While its maximum size is debated it is probably “only” between 6-7 meters (19.7 – 23.0 ft) in length. Rays can also grow quite large. The large, pelagic manta ray Manta birostris can grow at least 7 meters across (19.7 ft) and 2,721 kg (5,998.8 lbs) (McClain et al., 2015).
Elasmobranchs are found all around the world, mostly in marine habitats, although some ray species are found in freshwater (including the enormous stingray Urogymnus polylepis). The bull shark, Carcharhinus leucas, can tolerate freshwater for periods of time, and the sharks of the genus Glyphis can live permanently in freshwater. In marine habitats elasmobranchs inhabit all environments, with many rays and species such as the nurse shark inhabiting the benthos while other species, like the oceanic white-tipped shark and the blue shark being pelagic species of the open sea.
Elasmobranchs show a variety of reproductive modes. Some are oviparous, some are ovoviviparous, and some are viviparous (Wourms, 1977). In some shark species developing embryos feed on unfertilized eggs that the mother “lays” into the uterus, while in others the embryos eat their siblings. In sand tiger sharks (Carcharias taurus) multiple embryos begin developing in each of the two uterine horns, but the first embryo to become large enough eats all other embryos, leaving only two offspring (Chapman et al., 2013).
Feeding in elasmobranchs is highly variable, given the basic constraint that all are carnivores. Rays tend to be generalist carnivores on benthic animals, including hard-shelled ones. Some sharks follow this route, engaging in suction-feeding on smaller organisms (Wilga et al., 2007). The very largest modern shark species, the whale shark and the basking shark (Cetorhinus maximus) are suspension feeders. The cookie cutter shark (Isistius brasiliensis) is a small parasite that bites circular chunks of flesh from much larger animals and appears to use bioluminescence to attract these animals (Widder, 1998). Many sharks are active predators as well, eating a variety of actively-swimming prey.
One of the sharks best-known to the general public, the great white (Carcharodon carcharias), is actually not a particularly good example of a standard elasmobranch. Great white sharks belong to a small group of extremely fast-swimming sharks (the Lamnidae) that have reorganized their bodies to achieve higher speeds (Donley et al., 2004). At least some species in this group, including the great white, warm their bodies to temperatures higher than that of the surrounding water (Goldman, 1997). These are probably both adaptations to hunting fast, agile prey like seals and dolphins, prey that many other sharks would not be successful hunting.
Euteleostomi (1.0.0)
Traditionally the gnathostome fish have been split into two groups, the Chondrichthyes and the Osteichthyes (literally the cartilage-fish and the bone-fish). With the rise of phylogenetic taxonomy there is no longer a place for a bony fish group that does not include the tetrapods (land vertebrates). For some scientists this means that Osteichthyes now includes a number of non-fish. For others the term “Osteichthyes” is too tied to an old classification scheme and so a new term is needed. This text will use this new term, Euteleostomi, which unambiguously refers to the bony fish and the tetrapods. However, the term Osteichthyes remains in common use as well.
Euteleostomes are a diverse group in which some groups have secondarily lost almost any of the basic euteleostome characteristics one cares to name. However, three euteleostome characteristics are especially important for understanding this group.
The first is a skeleton made of bone. Bone is actually much more ancient than the origin of the eutelestomes, having originally evolved as a dermal element (Doherty et al., 2015). Many early fossil fish, including non-gnathostome fish, had bony armor, and it is likely that the Chondrichthyes have secondarily lost bone. Bony skeletons provide euteleostomes with two advantages. One is that bone stores calcium, an essential element for proper nerve conduction. The other is the high strength of bone, especially in compression.
The second is fins supported by fin-rays. All euteleostome fish retain this characteristic and these fin-rays have become toes in the vertebrates. The advantages of this fish structure are discussed in the section on the Actinopterygii.
The third characteristic is the most debated, but it seems probable that the earliest euteleostome had some sort of air-breathing organ (Perry et al., 2001; Longo et al., 2013). In modern tetrapods as well as several groups of fish this organ remains as the lungs. In many other euteleostomes this air-filled organ is the swim-bladder, which helps a swimming fish with buoyancy control.
Actinopterygii (1.0.0)
(Traditionally a Class, approximately 32,513 species, 48.66% of vertebrate species)
Actinopterygii is such a large group of vertebrates that it is probably fair to say that the “standard” vertebrate is an actinopterygian. 95% of all living fish species are actinopterygians and the range of diversity in this group is such that it is often hard to make general statements about the group that are true of all members.
Actinopterygians do have several major factors that differentiate them visibly from the chondrichthyans. First, their fins follow the euteleostome pattern with fin rays supporting a thin membrane between them. This means that actinopterygian fins can fold and “collapse”, allowing for a wide range of fin shapes since fin shapes can be altered “on the fly”. For instance, the flying fish (Exocoetidae) have long “wings” for pectoral fins that allow them to glide in the air but would interfere with swimming. These fins fold against the body, allowing the fish to have longer fins than it would be able to have otherwise.
Second, many actinopterygians are quite visibly scaly. Where chondrichthyan scales are small and visibly do little more than give the shark a roughened texture many actinopterygians sport large scales that may serve to improve the hydrodynamics of the fish or even armor it against predators. However, some actinopterygians do completely lack scales, include the catfishes, sturgeons, and some eels.
Thirdly, most actinopterygians have caudal fins in which both upper and lower lobes are the same size, a condition known as homocercal. Sturgeons are notable exceptions. The heterocercal tail of a shark is thought to provide lift, and since actinopterygians have swim bladders to perform this task the homocercal tail may be optimized for thrust instead.
Finally, unlike elasmobranchs, no actinopterygian has multiple gill slits. Instead, the gills are housed behind a structure called an operculum and all water is directed through the gill chamber and out from under the trailing edge of the operculum. The bones of the operculum are used to anchor muscles that allow actinopterygids to pump water out their gills more effectively.
A complete review of the basic actinopterygian clades is beyond the scope of this text but what follows is a discussion of some of the diversity represented within this clade. It is worth noting that most actinopterygians are teleosts, a group that excludes only the gars (Lepisosteiformes, 7 species), bowfin (Amiiformes, 1 species), bichirs/reedfish (Polypteriformes, 14 species), and sturgeons (Acipenseriformes, 27 species). This clade has a skull morphology that allows it to extend its upper jaw forward, making for more efficient suction-feeding.
Diversity of Size and Shape
The smallest actinopterygians are less than 10 mm in total body length, and many species, including many species popular as pets, are extremely small. However, actinopterygians also include extremely large species. Members of the family Molidae are hatched at extremely small sizes but grow to over a 1,000 kg, with the largest species (Mola mola) topping out at 2.3 tonnes (Roach, 2003). The oarfish (Regalecus glesne), an elongate ribbon-like fish, reaches 8 m (26.2 ft) in length (Roberts, 2012). In freshwater the Mekong giant catfish (Pangasianodon gigas) reaches 300 kg (770 lbs) in weight (Kottelat, 2001).
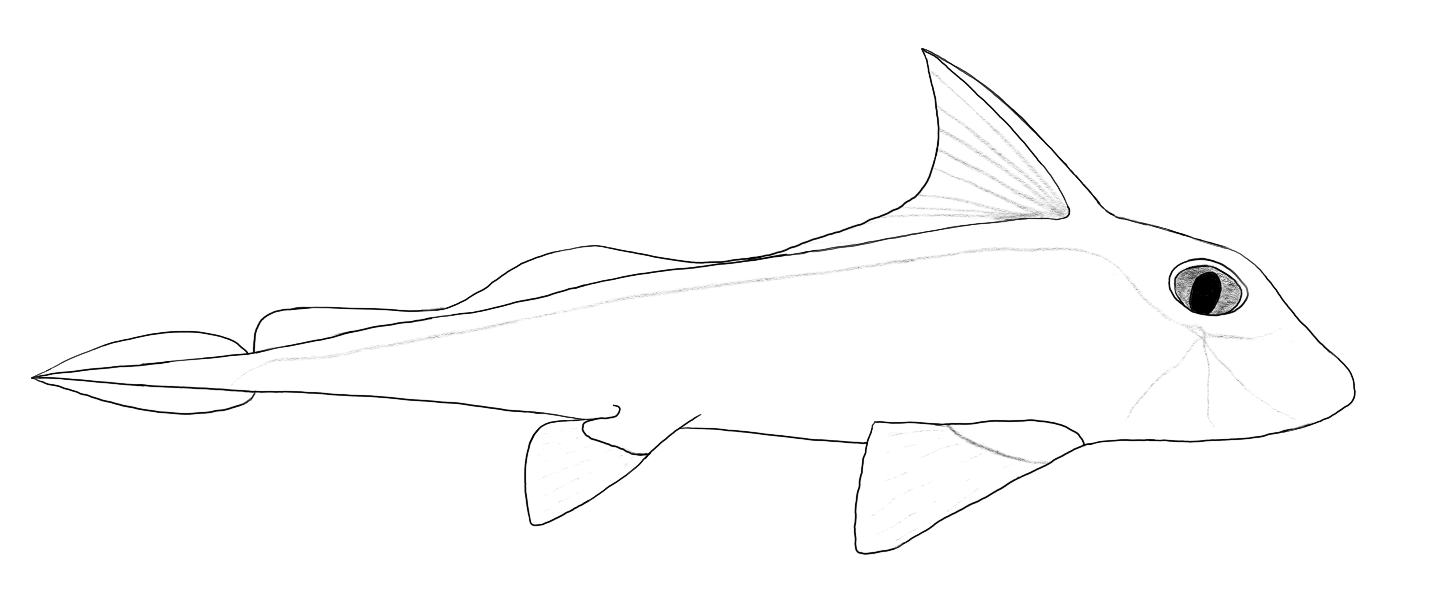
Shape also varies widely. The basic shape of an actinopterygian should be familiar from the fin diagram from the gnathostome section and is illustrated here by a red-tailed catfish, Phractocephalus hemioliopterus, a large, predatory catfish from the Amazon. All of the expected fins are present and the tail provides the main method of propulsion, although the whole body flexes to some degree during the swimming stroke.
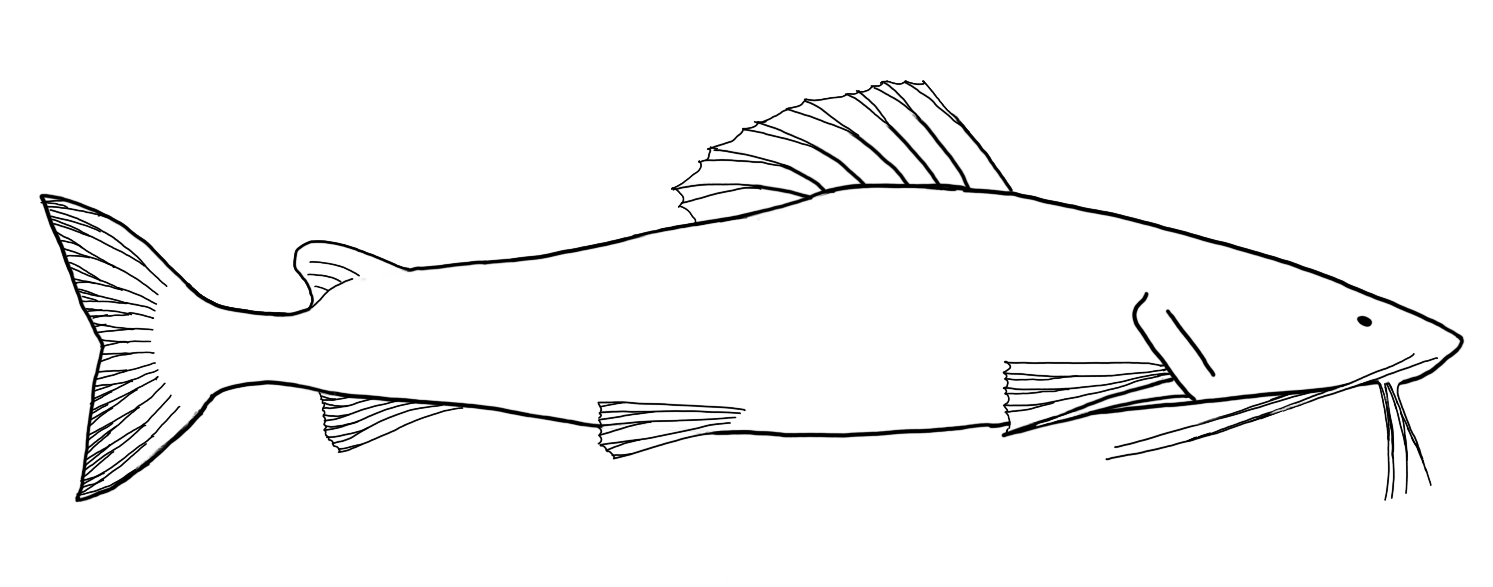
The previously-mentioned Mola mola presents a highly divergent morphology, in which the body is roughly circular with long, triangular fins (the first dorsal and anal fins) extended dorsally and ventrally from the body. This fish moves by “flapping” these fins as it cruises the open ocean. The tail is reduced to a knobby structure called a clavus which has no role in propulsion. The mouth is beaked and permanently open.

An opposite trend is seen in the green moray (Gymnothorax funebris) which is a long, snake-shaped fish. The dorsal, anal, and caudal fins have fused together and the paired fins are absent. Morays, and other eel-shaped fish, swim by sending an S-shaped wave down the entire body. This method of swimming is not particularly fast but it does provide a high degree of maneuverability and involves much of the eel’s body muscle making it efficient.
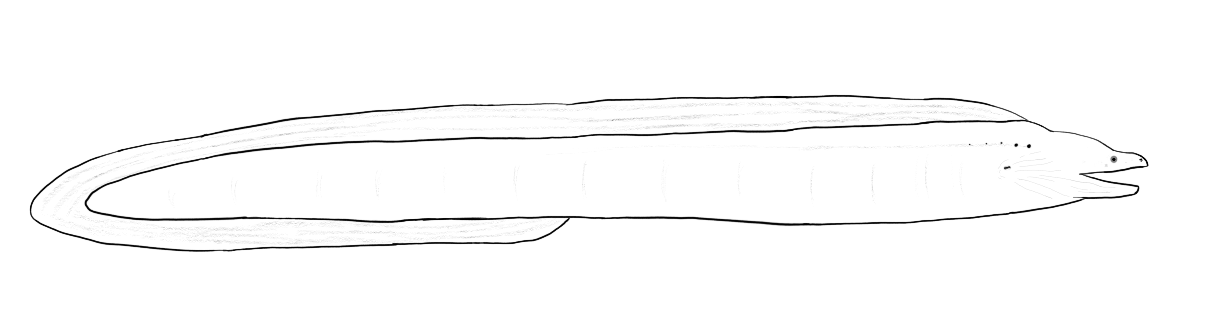
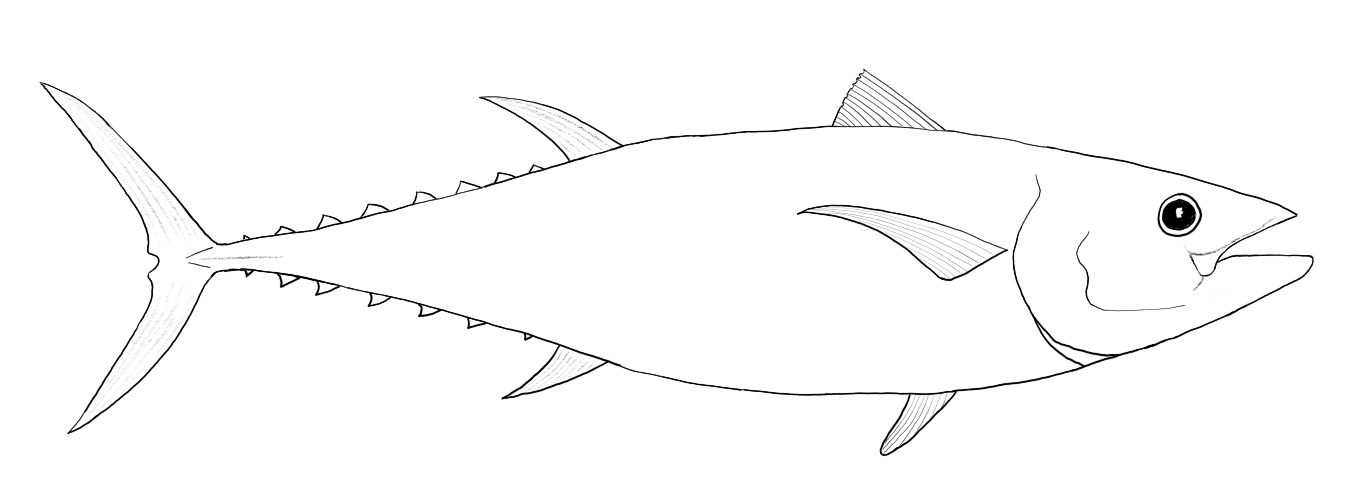
High speed swimming is seen in tuna (Thunnini) which hold their torpedo-shaped bodies stiff and pump their crescent-moon shaped tail (termed “semi-lunate”) back and forth at high speed. Most of a tuna’s fins have the same semi-lunate shape as the tail and there are a series of small finlets that run along the midline of the tail dorsally and ventrally. In addition to their body shape tuna are also capable of holding their body temperature above that of the surrounding water.
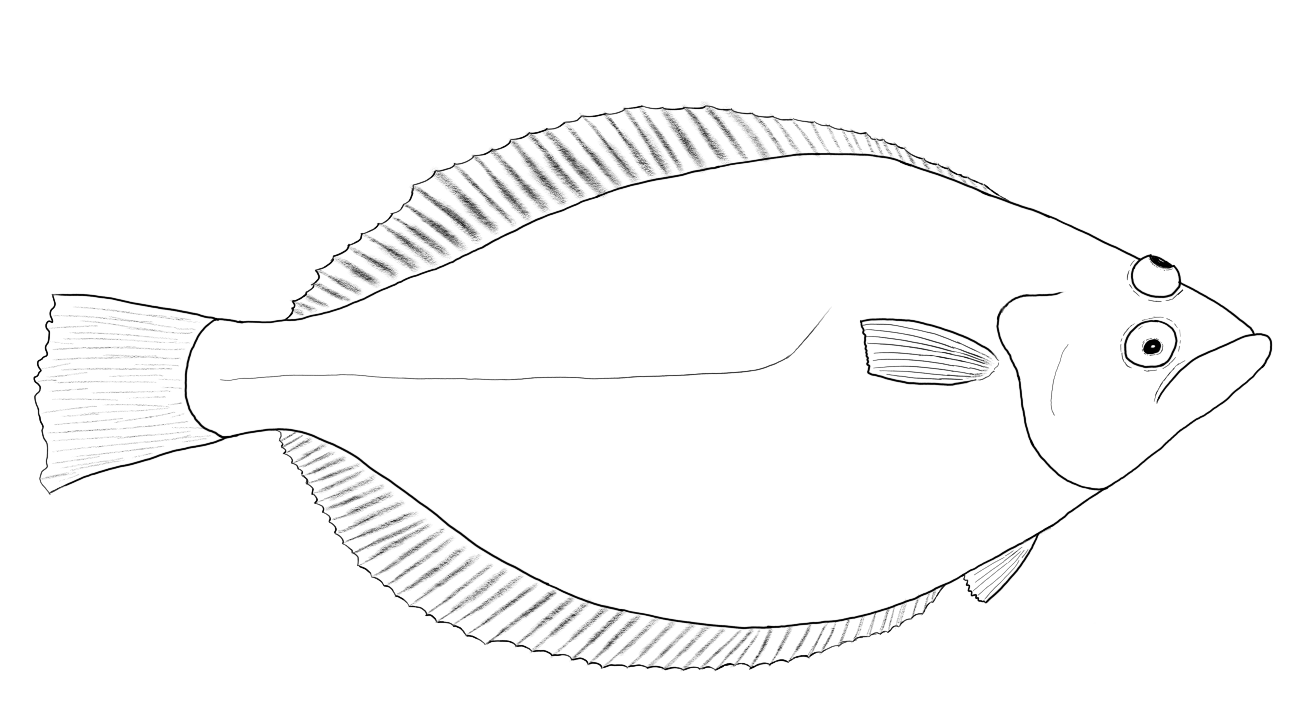
The ray-equivalent in actinopterygians is found in the flatfish (Pleuronectiformes) which are flattened, bottom-living species. Unlike rays which are dorsoventrally flattened flatfish are laterally flattened and live their lives on one side as adults (although they are born as “normal” fish). In the process of development the eye on one side of the body shifts position until it ends up positioned dorsally on the opposite side of the head.
Diversity of Habitat
Actinopterygians are found in almost all watery environments. In marine environments actinopterygians are found everywhere. Some species (like tuna) are pelagic zone specialists. Others, like many eels, are specialists of structured environments with many obstacles to navigate. Some actinopterygians live not only on the ocean floor (flatfish, scorpionfish) but in it, like garden eels and many gobies that burrow in sandy substrate in shallow water. In the deep ocean some actinopterygians are found with bioluminescent organs to attract prey (anglerfish). One group of actinopterygians, the tadpole-like snailfish (Liparidae), appear to include the deepest-living fish species. One species, Pseudoliparis swirei, has been described from 7.97 km (4.95 miles) deep (Gerringer et al., 2017) and a trip into the Marianas trench spotted what they believe to be a related species even deeper down. An entire clade of actinopterygians, the Notothenioidei, are cold-water specialists found around Antarctica.
Half of all actinopterygians are found in freshwater, an astonishing fact when one considers that freshwater makes up 0.0093% of the Earth’s surface and the oceans cover roughly two-thirds of the planet (Bone & Moore, 2008). Many fish clades are either marine or freshwater specialists. For instance, the Ostariophysi include several of the largest traditional families of fish, including the Siluriformes (catfish), Cypriniformes (carps, minnows, and loaches), and Characiformes (tetras, piranhas, and allies) nearly all of which are freshwater. The closest relatives of the Ostariophysi include the Clupeiformes, and almost entirely marine group.
Some fish also move between marine environments and freshwater. This is normally part of a migratory pattern based around reproduction, as in salmon that leave the ocean to spawn in freshwater.
Within freshwater environments actinopterygians include large-bodied species that require deep rivers or large lakes and also short-lived, small species that can inhabit bodies of water that dry out and refill regularly. For instance, alligator gar (Atractosteus spatula) can reach 2 meters (6.5 ft) and live up to 27 years, maturing around 5 years of age (Binion et al., 2015). The Amazon catfish Brachyplatystoma rousseauxii can reach similar lengths and breeds between its third and fourth year (Cordoba et al., 2013). On the other end of the spectrum is Nothobranchius furzeri, a fish that is around 3 cm (1.2 inches) long and lives only 4-6 months under optimal conditions, and can reproduce 14 days after hatching (Harel et al., 2015; Vrtilek et al, 2018). This fish lives in small bodies of water that dry up regularly, in as little as 3-5 weeks. When they do the fish’s eggs survive although the adults die.
One fundamental challenge to freshwater habitats is that they are often small bodies of water that can rapidly become deoxygenated under the right conditions. In response, many actinopterygians have evolved the ability to breathe air using a variety of organs. These include gar (which breathe air with their swimbladders), labyrinth fish (which include the popular betta fish and the snakeheads, which breathe air with an organ behind the gills), callichthyid catfish (which breathe air with their intestines), clariid catfish (which breathe air with an organ near the gills), bowfin (which breathe air with their swimbladders), the arapaima (which breathes air with its swimbladder), and the tarpons (which also breathes air with its swimbladder). This is not an all-inclusive list but it serves to demonstrate an important point: many freshwater actinopterygians, and a few marine ones (like the tarpon), can breath air.
One step beyond air-breathing is moving on land. Many species of air-breathing fish cannot move on land at all, including the gouramis and the arapaima. Others, like the clariid catfish, known as walking catfish, are famous for it. For the most part this is an occasional behavior used to move between bodies of water, but for one group, the mudskippers (Oxudercidae), most feeding and courtship is performed on land (although near the water).
Diversity of Diet
Actinopterygians show the same diversity of diets that they do in morphology. Probably the most common diet is a predatory diet. Most actinopterygians are suction-feeders, a style of feeding where the mouth is opened suddenly to create suction that draws in prey (Wilga et al., 2007). This means that many actinopterygians are also gape-limited predators, which means that the size prey that they can swallow is limited by the size of their mouths (their gape size). Some species, such as piranhas and barracudas, slice prey into sections to get around this limit, but most actinopterygians are predators of things that they can swallow whole. However, this has also led to some enormous mouth sizes. In groups like frogfish, groupers, and catfish an enormous mouth allows an actinopterygian to swallow correspondingly large prey items.
Suspension feeding is also common in actinopterygians, especially in marine forms. Many of the Clupeidae are suspension feeders and form an important part of the pelagic food web because of this. American paddlefish (Polyodon spathula) are an example of a (much larger) freshwater suspension-feeding actinopterygian.
Herbivory is the least common diet for actinopterygians. However, where it is practiced it is often a very successful strategy. The Loricariidae are the largest family of catfish and this diversity may be driven by the fact that many members are sucker-mouth fish that scrape algae from their habitats, allowing them to exploit a niche unavailable to many other fish. Surgeonfish and damselfish are marine herbivores.
Diversity in Reproduction
The basic mode of reproduction in actinopterygians is oviparity with external fertilization. In many oceanic species eggs hatch first into planktonic larvae that then become fry that then grow into adults. The giant ocean sunfish, Mola mola, is not only the largest actinopterygian but also the most prolific, laying up to 300 million eggs at a time (Tortonese, 1986). These eggs hatch into tiny larvae that progress through a series of stages which look little like the adult, before finally reaching a stage at which they are easily recognizable as Mola mola. Given the number of eggs per adult female juvenile mortality must be extremely high.
In freshwater actinopterygians the larval stage is normally bypassed, probably because freshwater environments always have one-way flow (from the river’s head to the river’s mouth, passing through any intermediate ponds and lakes) and so floating larvae would be continuously pushed towards the sea. Some marine actinopterygians also migrate into freshwater to spawn, including the well-known migrations of the various salmon species. In many salmon species the migrating fish are also genetically programmed to die at the end of their spawning run, having used the resources needed to keep them healthy for spawning. A few species migrate out of freshwater into the ocean, or brackish water, to spawn. These include the freshwater eels and some pimelodid catfish.
A few species of actinopterygians are internally fertilizing, and may have a viviparous or ovoviviparous mode of reproduction. The best-known of these are the live-bearers (Poeciliidae) of which several species (guppies and mollies) are commonly sold as pets.
Actinopterygians are also not all gonochoristic. Simultaneous hermaphroditism is rare, and self-fertilizing hermaphrodites are known only from the genus Kryptolebias. However, sequential hermaphroditism is much more common, in which an individual changes sex part way through life. In some species sex change is caused by body size, as in groupers, in which all individuals are born male and become female at a certain body size (Zho & Gui, 2010). In stoplight parrotfish, Sparisoma viride, this pattern is reversed, with small females becoming males as they grow, normally changing color as well (Cardwell & Liley, 1991). In other sequential hermaphrodites control over sex change is social, as in the well-known clownfish (Amphiprion melanopus) in which removal of the dominant female from a social group will cause the largest male to become female and take her place (Godwin & Thomas, 1993).
Parental care is also present in some actinopterygians. In some species it is limited to defense of a nest site and ends when the young hatch. In others the young are also guarded, as in convict cichlids (Amatitlania nigrofasciata) where both male and female guard the offspring until they reach a suitable size. In some cichlids, jawfishes, cardinalfishes, and sea catfishes the eggs and sometimes fry are carried in one of the parent’s mouths (in cichlids it is normally the female, in the other groups is it the male) to keep them safe. In a few species parental care goes further than this. In the genus Symphysodon, the discuses, the parent fish produce a nutritive mucus that the offspring feed from. This is known in discus in part because discus are valuable aquarium fish and great attention has been put into breeding them, but such adaptations may yet be discovered in less-well-investigated species as well
Sarcopterygii (1.0.0)
The Sarcopterygii was a group originally erected to encompass a group of mostly-extinct fish with fleshy fins (which is what the name means). However, sarcopterygian fish are the ancestors of the land vertebrates (tetrapods) and so the modern Sarcopterygii encompasses both the two groups of sarcopterygian fish and the tetrapods.
The major sarcopterygian feature is a fleshy “stalk” that the fin sits on. The the tetrapod this stalk became the limb with the fin rays as the digits.
Modern sarcopterygian fish represent only 8 species, around 0.01% of vertebrate species. While the sarcopterygian fish are not themselves a clade they will both be treated briefly here.
Actinistia
The Actinistia was once a large group encompassing many kinds of fish. Today the group is represented only by the coelacanths. Early coelacanths were probably reasonably morphologically diverse (Friedman & Coates, 2006) but today only two species remain, both in the genus Latimeria. The discovery of the extant coelacanths created major ripples in scientific circles as the entire clade of coelacanths had previously been supposed to have died out during the Mesozoic or in the extinction event that ended that era.
Modern coelacanths are deep-water, nocturnal fish (Fricke et al. 1991; Yokayama et al. 1999). They move largely by drifting and are large (1.5 m, 5 ft) carnivorous fish. They are ovoviviparous (Smith et al., 1975). The two species appear virtually identical but appear to have diverged some 40 million years ago (Inoue et al., 2005), suggesting that there are strong forces (either selective or genetic) that keep them in their current morphology.
Dipnoi
The Dipnoi are today represented by six species of lungfish. Lungfish have a fossil record extending into the Devonian and many of these fossils are recognizable as lungfish. However, Lloyd et al. (2012) showed that lungfish “stabilized” at their current form much later, around the end of the Cretaceous.
Lungfish are relatively large (all species can reach more than 1 meter in length) fish shaped like thick eels, but with sarcopterygian fins. All species are omnivores. Four of the modern species live in Africa, one in South America, and one (less related to the others) in Australia. All species have lungs and can breathe air, although the Australian species cannot live for extended periods out of water (Allen et al., 2002). In at least some of the other species access to air is required for the individual to survive (Babiker, 1979).
Most notably, the African and South American species respond to drought by burying themselves in the mud and entering a period of extended torpor. In this state they breathe air and their metabolism drops, allowing them to remain alive until the rains come.
Tetrapoda (1.0.0)
Prior to the evolution of the tetrapods the story of vertebrate life was a story of aquatic life. While some actinopterygian and sarcopterygian fish rose to the surface to gulp air or slithered across the ground to find a new river or pond the focus of vertebrate life was water.
For the earliest tetrapods dependence on land did not come quickly. The Devonian animal Ichthyostega shows many tetrapod features, including limbs that can bear weight on land and a reinforced rib cage that would allow it to crawl on land without placing undue pressure on its organs. However, Ichthyostega also shows a broad, finned tail that makes sense only in the context of swimming. It appears that many early tetrapods moved over land either to find water or used their limbs to maneuver in shallow water (and advantage for a large predator hunting small fish).
The shift on to land required dealing with many new challenges. Aquatic vertebrates always have access to water, both to drink and to flush away waste. Terrestrial vertebrates needed to find drinking water, retain it, and not waste it flushing waste products from their bodies. The entire process of excreting waste had to change, as is many fish many waste products leak directly out of the gills.
Terrestrial vertebrates also had to deal much more directly with the problem of gravity. Aquatic vertebrates can obtain neutral buoyancy and float their bodies but terrestrial vertebrates must support their bodies against gravity. This is especially important for animals which need to get their ventral surfaces clear of the ground for faster movement.
Air and water also pass sound and light waves differently than does water. Ears and eyes that worked well under water would have worked poorly for animals venturing onto land, and fundamental senses like the lateral line (which detect water motion) and electroreception (which detects the electrical fields of animals in water) have no terrestrial equivalent.
However, the tetrapod experiment has been hugely successful, and today around half of all vertebrates belong to this lineage, including some who have secondarily returned to the water
Amphibia (1.0.0)
(Traditionally a class, approximately 6,489 species, 9.64% of vertebrate species)
The modern Amphibia (Lissamphibia) are sometimes depicted as holdovers from an ancient era – tetrapods that “forgot” to get with the program and finish the tetrapod march towards land dominance. Nothing could be further from the truth. Amphibians are highly-derived animals that have evolved to fit their (amphibious) lifestyle.
One notable point of divergence between amphibians and both sarcopterygian fish and the first amniotes is that amphibians lack scales. The more terrestrial amphibians sometimes have a more keratinized skin, but the majority of amphibians have a smooth, think skin covered in a layer of mucus. This mucus often contains toxins, ranging from poisons that give the animal a foul taste (many of the toads common to the United States) to toxins that can kill large vertebrate predators in very small doses (the Dendrobatidae, the poison dart frogs). Poisonous amphibians are often brightly colored to ward off predators. The thin skin of amphibians allows them to take oxygen up directly through their skins (cutaneous respiration), an ability that can be used to respire in both air and water. For some amphibians cutaneous respiration is the only important form of respiration. The Plethodontidae are a group of salamanders, most of whom live on land, who are known as the lungless salamanders because they lack lungs and breathe entirely through their skins. The Cryptobranchidae are aquatic salamanders without gills who do not display surfacing behavior to breathe with their extremely small lungs, and instead rely on oxygen they take up through their heavily-folded skins.
Amphibians are also characterized by having a gelatinous egg (without a shell) and, often, a larval stage. These larvae are normally fully aquatic and possess gills, although variations on this theme exist. The anuran Indirana semipalmata has terrestrial tadpoles that stay wet from rain (Gaitonde & Giri, 2014), dendrobatids often deposit tadpoles in the standing water in bromeliads (Weygoldt, 1987), in Darwin’s frog (Rhinoderma darwinii) tadpoles develop inside their father’s vocal sack and leave his mouth after metamorphosis, and Leptodactylus pentadactylus frogs lay eggs in foam nests which either flood, releasing the tadpoles, or hatch out where the tadpoles live in the foam eating the other eggs (Muedeking & Heyer, 1976). Direct development (no larval stage) is also known in many more terrestrial species. Some amphibians in all three groups are viviparous, with this strategy being most widespread in the Gymnophiona (Xavier, 1977; Buckley et al., 2007; Gower et al., 2008).
All three groups of amphibians possess members that are wholly aquatic and some members that are terrestrial to the extent that they probably never enter bodies of water. However, the vast majority of amphibians are either species that inhabit the edges of bodies of water or live in other wet habitats.
Anura
By total species numbers most amphibians are anurans, at around 5,685 species. Anurans are composed of frogs (any smooth-skinned anuran) and toads (any bumpy-skinned anuran). The basic anuran body plan is extremely heavily modified from the tetrapod norm with notable changes including the reduction of the ribs, the loss of a tail in adults, fusion of many vertebrae to form the urostyle, and the elongation of hindlimbs (Přikryl et al., 2009). While this body form is strongly associated with jumping by many people anurans also involve a large number of independently-evolved “tree frogs” which climb, burrowing members, and fully-aquatic swimming frogs. The “toads” are also independently-evolved, as the term refers to any anuran with keratinized, bumpy skin, a common adaptation to prevent water-loss.
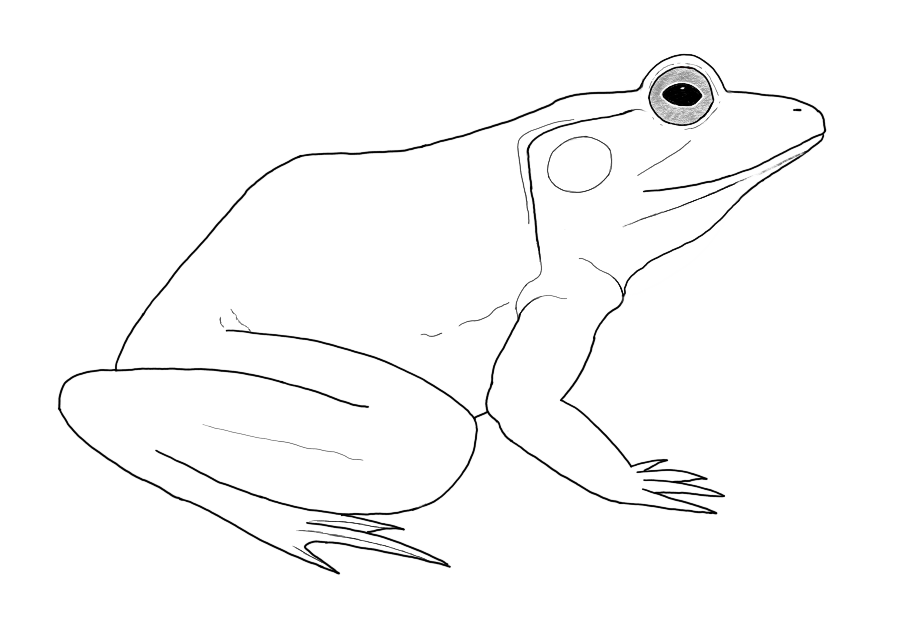
Adult anurans are carnivores, and many are indiscriminate about their prey, shoving any animal small enough into their mouths. African bullfrogs (Pyxicephalus adspersus) have been recorded eating young cobras and a fledgling chicken (Branch, 1976) while the large Surinam horned frog (Ceratophrys cornuta) has been recorded eating rodents, lizards, other frogs, and snakes (Chavez et al., 2011). While frogs are often said to use their tongues to catch prey this is only true of some anurans and only for smaller prey. The tongue is attached at the front of the mouth, allowing it to be projected further out of the mouth than might be expected. In other cases the anuran will use its forefeet to push prey into its mouth where small teeth can hold it. The eyes of anurans can also drop through the roof of the skull, producing bulges on the roof of the mouth that can help shove prey down the anuran’s throat.
In contrast, many anuran larvae (tadpoles) are herbivores, although carnivorous and suspension feeding species also exist. In some species individual tadpoles may be either carnivores or herbivores, as in the spadefoot toads (genus Scaphiopus) where a single egg mass will hatch out into quick-developing, cannibalistic, carnivorous tadpoles and slow-developing, more numerous, omnivorous tadpoles (Pfennig, 1992).
Anurans also undergo the most dramatic metamorphosis of all the amphibians, as their larvae change the position and structure of the mouth, lose the tail, grow limbs, move the eyes, switch from gills to lungs, and, in many species, change their digestive tract to accommodate a carnivorous diet.
Anurans show a wide diversity of reproductive strategies, many of which involve males calling for females. Most anurans lay eggs but the location of the eggs and the amount of parental care varies widely. In some species, such as the midwife toad, one parent carries the eggs. In the Surinam toad (a very flat aquatic anuran) the male places the fertilized eggs on the female’s back and she allows them to sink into a thickened area of skin. The eggs hatch, complete their development, and tiny toadlets burst out of their mother’s back.
Caudata
Caudata (or Urodela) are the salamanders and newts. While they compose only 9% of living amphibian species they can be incredibly abundant in some ecosystems. In a study in New Hampshire, USA, the biomass of salamanders was twice that of birds and as large as that of small mammals (Burton & Likens, 1975). 93.5% of this was a single species, Plethodon cinereus.
Caudates are called caudates because they, unlike anurans, have tails. In fact, salamanders closely resemble small lizards except that they lack scales and claws. It is generally believed that the ancestral amphibian resembled a caudate and that the other two groups represent divergence from this form.
Divergence from the basic form in caudates is relatively common. Caudate larvae resemble adult caudates much more than anuran larvae resemble adult anurans, and many caudates can become reproductively mature while remaining in a larval body form. This neotenous morphology resembles an adult salamander except that it has external gills, like the larvae. In some species this form is temporary and the salamander can metamorphose into a “normal” adult if conditions change, but in other species, including the extremely-well-studied axolotl (Ambystoma mexicanum) adults remain in the neotenous form throughout their lives.
Caudates also have several partially-limbless species. Both sirens (Sirenidae) and amphiumas (Amphiumidae) lack one set of limbs, the back ones in sirens and the front ones in amphiumas. These eel-like salamanders are fully aquatic species but can use their bodies to both swim and crawl in swampy habitats. Both clades are found only in North America, which is a hotspot for caudate diversity.
Caudates also vary markedly in size. Many plethodontids, one of the most diverse clades, are smaller than a person’s little finger (in part because plethodontids rely entirely on cutaneous respiration and must stay small for this to work). Cryptobranchids, on the other hand, are all large. The smallest species, the hellbender of the United States (Cryptobranchus alleganiensis) can grow to 70 cm (27.6 in) and the largest species, the Chinese giant salamander (Andrias davidianus – although there are probably multiple species lumped under this name) can reach 1.8 m (5.9 ft).
Caudates are generally predators. Large species, like the cryptobranchids, may eat fish, other amphibians, or other small vertebrates. Smaller species eat arthropods. In some of the plethodontids the tongue can be projected at high speed from the mouth, much as the tongue of a frog is often imagined to do.
Gymnophiona
The Gymnophiona, or caecilians, are a poorly-known group of limbless, burrowing amphibians found in tropical areas of the world. Some species are aquatic, and the rubber eel sometimes sold in fish stores is the caecilian Typhlonectes natans, although it is also able to live in damp soil. In general, caecilians look like very large worms with distinct, jawed heads, and very small tentacles near the nostrils.
Like all almost amphibians, caecilians are carnivores. The diets of caecilians are as poorly-reported as the rest of their biology but it appears to consist of any appropriately-sized animal. Records include many arthropods, earthworms, anuran eggs, larvae, and adults, fish, lizards, and snakes (Verdade et al., 2000; Presswell et al., 2002). Bite forces in at least some caecilians are quite high given their prey and are enhanced further by long-axis rotation (“death rolling”) after the prey has been bitten (Measey & Herrel, 2006).
Caecilians are best known for a high incidence of ovoviviparity and viviparity, and for their parental care. While caecilians are not all good parents several species engage in a behavior in which the mother generates nutritious extra layers of skin that her offspring then scrape off of her and eat (Kupfer et al., 2006).
Amniota (1.0.0)
One of the major barriers that held vertebrates back from fully utilizing the land was reproduction. In the water fertilization can be external (because sperm can swim through water) and the egg does not need to be protected from drying up. Out of water an egg can dry up and sperm cannot swim.
The amniotes are the group of tetrapods that solved this problem. Fertilization is internal in the amniotes, solving the problem of swimming sperm, and eggs have shells, preventing desiccation. However, adding a shell to an egg creates problems. A gelatinous fish egg brings oxygen in from outside and releases carbon dioxide and metabolic waste to the outside environment. A shelled egg cannot do this effectively, and so an amniotic egg also includes a series of membranes to manage these problems. These membranes remain in viviparous amniotes as uterine structures.
The basic membranes are: the yolk sac, which contains the yolk, the allantois, which holds nitrogenous waste from the developing embryo (and forms the umbilical cord in mammals), the amnion, which contains the embryo and fluid for the embryo, and the chorion, which is involved in gas exchange. The shell of the egg forms outside all of these membranes.
The amniotic egg allows amniotes to reproduce fully on land. Unsurprisingly, amniotes also show direct development, without larval stages.
The Diapsid-Synapsid Split (1.0.0)
Shortly after the evolution of the amniotes this clade split in two. One lineage, the diapsids, is today composed of the reptiles and birds. The other, the synapsids, is represented today only by the mammals.
The terms “synapsid” and “diapsid” originally referred directly to skull morphology. In amniote skulls there may be openings called temporal fenestra posterior to the eye orbit. Diapsids have two of these, which are often clearly visible in the skulls of many non-avian dinosaurs. Synapsids have one temporal fenestra. A lack of any temporal fenestra is called anapsid.
However, animals can also change their temporal fenestra over evolutionary time. Turtles have anapsid skulls and mammals effectively do as well, as their single temporal fenestra has been occluded by the sphenoid bone. Modern taxonomy follows phylogenetic principles and so the actual condition of the skull does not matter. Since turtles are descended from diapsids they are part of Diapsida, and, similarly, mammals are part of Synapsida. Somewhat confusingly, the name for a member of Diapsida is “diapsid”, exactly like the skull terminology, and the same issue occurs with “synapsid”. Be aware of the context for the words “synapsid” and “diapsid”.
Diapsida (1.0.0)
The modern diapsids are the reptiles and birds. This group of animals may also be called Reptilia, Eureptilia, Sauropsida, or Sauria. While Reptilia is probably an inadvisable term to use, since it has long baggage as a paraphyletic taxon (it did not traditionally include birds), the other terms all mean the same thing when restricted to living species. Specifically, all living reptiles and birds fit within Sauria, which fits within Diapsida, which fits within Eureptilia, which fits within Sauropsida.
The relationships of the modern diapsids are, with one exception, now generally agreed upon. The Lepidosauria includes the tuataras and the lizards. Snakes are now understood to be a large, limbless clade of lizards. Another group, the archosaurs, includes the crocodylians and the dinosaurs. Birds are living dinosaurs and fit within this group. The position of the Testudines, the turtles and tortoises, has been highly contested. Originally placed outside of the diapsids altogether modern molecular phylogenies have placed them firmly within the diapsids. More recently studies have indicated that the Testudines are most closely related to the archosaurs (Crawford et al., 2012).
Within this group, the traditional reptiles (i.e., extant diapsids excluding birds) have some 10,233 species, approximately 15% of all vertebrate species.
Lepidosauria
The Lepidosauria include the Rhynchocephalia (tuataras) and Squamata (lizards and snakes).
Rhyncocephalia (traditionally an Order, 1 species, 0.01% of vertebrate species)
The Rhyncocephalia are currently restricted to one species, the tuatara of New Zealand. The tuatara is externally lizard-like, resembling an iguana in some aspects. Internally tuataras differ from lizards in many anatomical details, and in some cases appear to retain older characteristics that squamate reptiles have lost or modified.
The tuatara is a long-lived, slow-growing animal whose existence is threatened by non-native rats that eat eggs and possibly juveniles (Crook, 1973). In the wild females nest once every four years on average (Cree et al., 1992). This may be an adaptation to New Zealand’s climate, but it makes these reptiles vulnerable to factors that impact the success of their rare reproductive efforts.
Tuataras are carnivores, feeding mainly on large insects but also taking eggs and chicks of nesting petrels during the petrel breeding season as well as eating carrion (Walls, 1981). Walls described the tuatara’s feeding preferences as being those of a generalist carnivore, and stated that it would eat any animal of the right size that lacked sufficient deterrents.
Squamata (traditionally an Order, approximately 9,863 species, 96.38% of “reptile” species)
The Squamata is the largest reptilian diapsid clade, exceeded in size amongst the diapsids only by the birds. The squamates encompass both lizards and snakes. Lizards have evolved leglessness multiple times and it is now recognized that snakes represent only the most successful lineage of legless lizards and not an independently-evolved squamate clade. (For the purposes of this text “lizard” will be used as a non-phylogenetic term for non-snake squamates.) Approximately 60% of squamates are lizards and 40% are snakes.
All squamates possess a level of skull mobility that is unusual in an amniote. These kinetic skulls allow squamates to swallow surprisingly large food items.
Within the lizards body morphology varies considerably. The smallest lizards are, depending on how one measures, either the dwarf gecko (Sphaerodactylus ariasae) which has a maximum recorded snout-vent length of 18 mm (0.7 in) and a total length (i.e., adding the tail to the snout-vent length) of > 30 mm (> 1.18 in) or a tiny chameleon, Brookesia micra, with a 19.9 mm (0.78 in) maximum snout-vent length and a total length of only 29 mm (1.14 in) (Glaw et al., 2012). The largest lizard in the world is the Komodo dragon (Varanus komodoensis) which has been reliably measured at 3 m (9.8 ft) in total body length (Jessop et al., 2006). These species also vary considerably in shape. Geckos tend to be arboreal and have large feet with expanded toe pads. They are often shorter-bodied lizards with fat tails. Varanids, including the Komodo dragon, are often long, lean lizards that stand tall when walking. Chameleons are another familiar example of an extreme morphology in lizards, with laterally-compressed bodies, a tail that can curl downwards to grasp branches, toes that are fused to create two large toe-units, independently aimable eyes in cone-shaped turrets, often horns for display, and tongues that can be projected almost the length of the body to capture prey.

Leglessness is also common in squamates. While snakes are the best-known legless clade (Serpentes) a number of other “lizard” clades also have evolved legless sub-clades. The unusual amphisbaenians are a legless clade that was once thought to be distinct from the lizard-snake clade, but is now believed to be buried deep within the phylogeny of the other squamates (Vidal & Hedges, 1999; Townsend et al., 2004). The legless reptiles are burrowers that are generalist carnivores, eating many arthropods and occasional small vertebrates (Colli & Zamboni, 1999; Martín et al., 2013). Anguid lizards also include a number of legless forms, including the glass lizards and slow worms found in North America and Europe, and the Anniellidae, related to the anguids, are legless. The Pygopodidae are a legless (or mostly legless – the hind limbs are flaps) clade of geckos, some of which eat lizards (Patchell & Shine, 1986). The poorly-known and mostly-legless (males retain flap-like hindlimbs) Dibamidae appears to be the sister group to all other squamates. Both Scincidae (the skinks) and the spectacled lizards (Gymnophthalmidae) have many species with reduced limbs or no limbs at all.
Snakes themselves differ from other limbless squamates in several key characteristics. Snakes lack external ear openings, seen in many lizards, and the belly scales of most snakes are so wide that only one row is present on the venter. In external morphology snakes vary primarily in length and body width. The smallest snake in the world is the Lesser Antillean threadsnake (Tetracheilostoma carlae), with a total length of 100 mm (3.9 inches) (Glaw et al., 2012). The longest snake in the world is widely believed to be the reticulated python (Python reticulatus), although many large pythons and boas are close in size and the largest individuals of many species may have been hunted out of existence. The largest reliably-measured reticulated python was 6.95 m (22.8 ft) (Fredriksson et al., 2005). The green anaconda (Eunectes murinus) is the heaviest snake species since green anacondas approach reticulated python lengths but are extremely heavy-bodied snakes (perhaps because they are semi-aquatic) and weigh much more than reticulated pythons of the same length. Constricting snakes are normally much more heavily built than their non-constricting counterparts.

A small number of snakes live most of their lives at sea. These snakes often have a laterally-compressed tail that is slightly leaf-shaped. They often have even-sized scales across their venters as they do not crawl on land.
The diet of squamates is generally carnivorous. In lizards herbivory and omnivory are more common, but all snakes, with the exception of some specialist egg-eaters, are predators. Predatory lizards may capture their prey with extensible tongues (as in chameleons), by grabbing prey and swallowing it whole, or by slicing it into pieces with sharp teeth. Some lizards have adaptations for crushing the shells of snails as well. Smaller lizards feed largely on arthropods but larger species may feed on vertebrates. The largest lizard, the Komodo dragon, hunts prey up to the size of water buffalo and deer using venom (Fry et al., 2009). The two species of Heloderma (the Gila monster and the beaded lizard) are also venomous.
Many largely-carnivorous lizards are actually omnivores, feeding on easily-digestible plant parts such as fruit. A switch to a herbivorous diet has occurred in a number of species, including some monitors (Varanidae), a group that is known for its active predatory behavior (Law et al., 2016). The iguanas are the best known herbivores (Troyer, 1984) and this group includes the unusual marine iguana (Amblyrhynchus cristatus) that dives to forage on algae in the ocean before hauling itself on to the shore to warm up and digest (Wikelski et al., 1993).
Amongst predatory snake species predation is divided between three major modes of prey capture. Some snakes, especially snakes that feed on very small prey, like insects, simply swallow their prey without killing it. Other snakes are constrictors which coil around their prey and apply extreme pressure to the thorax. The mechanisms by which constriction kills prey are unclear and may include preventing the prey from inhaling, stopping the heart, dislocating the spine, or forcing blood into the brain at high pressure (Penning et al., 2015). However constriction works it is effective. Large constrictors have been known to eat animals such as deer, pigs, small bear species, and occasionally humans (Fredriksson et al., 2005; Boback et al., 2016). Constricting snakes often rely on ambush to attack their prey and are equipped with many backwards-pointing teeth to grab prey as they constrict it. Venom is the last method by which snakes subdue prey. Many kinds of venoms are present in snakes and many snakes are able to rapidly disable prey with venom. In most venomous snakes the teeth are reduced, leaving a small number of long fangs, some of which fold back when not in use. Most venomous snakes use their venom defensively as well as offensively. When hunting, most venomous snakes bite and release, allowing a struggling animal to escape (rather than fight the snake) and die later. The snake then tracks the prey animal down and eats it.
All snakes face the issue of swallowing prey without being able to dismember it. Highly flexible skulls aid in this, but many snakes also target species that are easy to swallow, such as fish or other snakes. The largest venomous snake in the world, the king cobra (Ophiophagus hannah) bears a genus name that means “snake eater”, and many other snakes are specialized predators of other snake species.
Squamates are notable for having paired penises, called hemipenes. However, squamates are also notable for having a number of taxa in which no males exist and females give birth to female offspring that are born from unfertilized ova (parthenogenesis). These species appear to come about as a result of hybridization (Kearney et al., 2009) but facultative parthenogenesis, in which a species that normally reproduces sexually reproduces parthenogenetically some of the time, occurs also (Booth et al., 2012). Squamate reproduction is normally oviparous with no parental care but exceptions exist. Species of both lizards and snakes may be ovoviviparous or viviparous and parental care includes egg-guarding, egg-warming (some python species “shiver” to heat their eggs), and guarding offspring (Greene et a;, 2006; Lourdais et al., 2007).
Testudines (traditionally an Order, approximately 334 species, 3.36% of reptile species)
The turtles, tortoises, and terrapins are contained within the Testudines (also known as Chelonia or Chelonii). These names are used for different body shapes of testudines, do not correlate to phylogeny (i.e., not all tortoises are related), and are used differently in the USA and the UK.
Testudines have been very hard to place within the Diapsida in large part because of how different they are from other diapsids (their skull is actually anapsid, anatomically). The most dramatic of these differences is the presence of the shell, which is divided into the upper shell (carapace) and lower shell (plastron), which are joined together between the front and hind limb. Parts of the vertebral column and ribs are fused into the shell. Contrary to popular belief, a turtle can leave its shell only by being dismembered. Testudines also lack teeth, having a beak instead. Testudines tend to be slower animals but their shells prevent larger individuals from being preyed upon by many predators. One result of this is that testudines tend to be skewed towards a slow-maturing, long-life strategy. One study found that the relatively small yellow-bellied slider (Pseudemys scripta) took an average of eight years to mature (Gibbons et al., 1981) while another study found that the much larger loggerhead sea turtle (Caretta caretta) took an average of 45 years to mature, longer than the entire lifespan of many mammals (Scott et al., 2012). Many tortoise species are reliably judged to have lived longer than a century.
Testudines exhibit three basic body forms: the terrestrial form, in which the legs are pillar-like and the animal walks high off the ground, the freshwater form, in which the legs are held to the side, the feet splay out for swimming, and the plastron drags on the ground when on land, and the sea turtle, in which the legs are modified into flippers. Of these, the fully terrestrial forms have produced some extremely large species, including the Galapagos giant tortoise (genus Chelonoidis), which can weigh 400 kg (881.8 lbs). Sea turtles also tend to be large. The largest, the leatherback sea turtle (Dermochelys coriacea) can weigh 650 kg (1,433 lbs) (McClain et al., 2015). Leatherback sea turtles are also unusual in having a leathery, not hard shell (a trait shared with various soft-shelled freshwater turtles), and in being able to maintain its body at a higher temperature than the surrounding sea water (James & Mrosovsky, 2004).
Testudines eat more plant matter than most other reptiles. In some species plant matter is the primary diet, as in many terrestrial forms (Neira & Johnson, 1985). Galapagos tortoises may be important for seed dispersal on the Galapagos (Blake et al., 2012), which was not home to other large herbivores before humans arrived. In other testudines, including some sea turtles, plant matter becomes more important as the individual grows (Hart, 1983; Arthur et al., 2008). Even in testudines well-adapted for predation, such as the common snapping turtle (Chelydra serpentina), plant matter can be part of the diet (Punzo, 1975). However, snapping turtles also demonstrate another testudine dietary mode: use of the beak as a predatory tool. Snapping turtles are known to eat fish, amphibians, and insects (Punzo, 1975; Moldowan et al., 2015). No testudine is known to hunt prey that is large in comparison to its body size.
Testudines show no parental care in any species. Eggs are laid in nests dug into the substrate and abandoned. Temperature-dependent sex determination, in which the sex of the developing embryo is determined by the temperature it experiences as it grows, is relatively common in testudines (Bull & Vogt, 1979).
Archosauria
The archosaurs (a name which means “ruling reptiles”) are currently composed of the crocodylians and the birds. During the Mesozoic this group also included a large number of non-avian dinosaurs and the flying pterosaurs. Archosaurs have thecodont (socketed) teeth in forms with teeth, antorbital fenestra (a hold through the skull wall anterior to the eye), and mandibular fenestra (a hole in the jaw). Other uniting characteristics are less obvious to an untrained observer.
Modern phylogenies have placed the testudines nearer the archosaurs than to the Lepidosauria.
Crocodylians (traditionally an Order, approximately 25 species, 0.24% of reptile species)
Crocodylians, sometimes spelled crocodilians, are often depicted as living fossils, unchanged from the time of the dinosaurs. This is both true and false. It’s true in that the clade that includes the modern crocodylians split from other clades prior to the Mesozoic and it’s true in that crocodylians so similar to modern forms to have been grouped into the same genera have been present since at least the Eocene (Whiting & Hastings, 2014). It’s false in that modern crocodylians trace their origins, separate from other crocodylian-lineage animals, back only to the Cretaceous, making them younger than birds (Buscalioni et al., 1997; Salisbury et al., 2006).
All modern crocodylians are semi-aquatic predators with long jaws, short legs, long tails that are laterally compressed to aid in swimming, and with eyes and nostrils located on the top of the skull, allowing the animal to see and breathe without placing most of its body above the water. Crocodylians also have non-overlapping scales, heavily-cracked skin that creates the impression of facial scales, and scutes, bony projections covered by skin.
Crocodylians are also all large reptiles. The smallest crocodylian species is the dwarf caiman (Paleosuchus palpebro) which reaches 2.1 m (6.9 ft) in maximum length (Sanaiotti et al., 2010). The largest crocodylian is the saltwater crocodile, Crocodylus porosus. The largest reliably-measured individual was captured in the Philippines in 2011 as part of an attempt to capture a crocodile responsible for the deaths of two people. This individual was 6.17 m (20.2 ft) in length and estimates from this individual suggest that a skull preserved without a body represents an individual almost 7 m (23 ft) in length (Britton et al., 2012).
Crocodylians spend much of their time in water and move in water much more effectively than they do on land. In the water they propel themselves by undulating their tails with their limbs held against their sides, used only for steering. On land they may drag themselves forward or stand up in a “high walk” which allows them to move much more quickly, at the cost of the energy required to keep themselves held higher off the ground.
Crocodylians are split into three main lineages, the Crocodylidae (crocodiles), the Alligatoridae (caimans and alligators), and the Gavialidae (gharials, which have long, thin jaws). The Alligatoridae generally have jaws that taper less towards the rostral end than the Crocodylidae, giving alligators and caimans snouts with a “U” shaped end rather than a “V” shaped end.
All crocodylians are predators, although fruit-eating has recently been documented and may be more common than previously believed (Platt et al., 2013). What, exactly, crocodylians eat depends on their size and species. While most people are familiar with large crocodylians hunting terrestrial prey, including humans, by lunging from the water many species of crocodylians do most of their hunting in the water (Magnusson et al., 1987; Bhattarai, 2015). Nile crocodiles (Crocodylus niloticus) have been shown to switch from arthropods to a mixed diet of fish, crustaceans, and amphibians, to a mostly fish diet as they grew (Wallace & Leslie, 2008), although this study did not examine large adults, who are known to take large terrestrial mammals as prey. While crocodylians preying on large prey is dramatic and well-known a study on American alligators (Alligator mississippiensis) in Florida found that adults ate mostly smaller animals, including large fish, turtles, rabbits, and muskrats (Delany & Abercrombie, 1986). The extreme bite forces produced by crocodylians allow them to subdue large prey as well as break bony armor. In Australia large adult saltwater crocodiles (Crocodylus porosus) have been observed killing and eating adult sea turtles, breaking through the shell to consume the turtle (Whiting & Whiting, 2011). In some populations saltwater crocodiles will actually follow nesting turtles onto beaches and attack them on land.
While crocodylians are frequently considered to be primitive and stupid recent work has shown that at least two species of crocodylians have learned to use sticks to lure birds close enough to capture (Dinets et al., 2015). This behavior takes advantage of the nesting behavior of some bird species, in which nesting adults will bring appropriate sticks back to the nest site to build nests. However, as this behavior occurs only at certain times of year the crocodylians’ mastery of this strategy requires crocodylians to learn a hunting strategy that is also time-dependent.
Crocodylians show parental care, with the female building and guarding a nest. In many species the female will also guard the offspring. The nest is normally built from rotting vegetation that generates heat as it rots, warming the eggs. Temperature-dependent sex determination is present in at least some crocodylians, making nest temperatures critical (Lang & Andrews, 1994).
Aves (traditionally an Order, approximately 10,357 species, 15.50% of vertebrate species)
Sometime in the Triassic an archosaur appeared that would define the rest of the Mesozoic: the first dinosaur. Dinosaurs would go on to become the largest land animals ever and to fill a variety of ecological roles, but it is generally unrecognized by the public that feathers appeared quite early in dinosaur evolution, perhaps even prior to the first true dinosaur (Xu et al., 2009). Feathers are well-known from the dinosaur clade Theropoda, the clade composed of bipedal predators, including well-known dinosaurs such as Velociraptor mongoliensis and Tyrannosaurus rex. Starting with the discovery of Archaeopteryx lithographica in 1861 and continuing on with more recent well-preserved fossils in China the transition between small, feathered, non-flying dinosaurs and small, feathered, flying dinosaurs (the birds) has become clear.
Modern birds fall within the Aves and all members of the group are highly modified by ancestral flight, whether or not the modern species flies. Non-bird theropods used their forelimbs to grasp prey and perhaps climb, but birds have lost this functionality completely, and have wings for forelimbs instead. In the recently-extinct, flightless moas the forelimbs disappeared rather than re-evolve their original function because modifications to the forelimbs are too extreme to be easily reversed over evolutionary time. The means that all birds are bipeds and the wings are used in flight, underwater swimming, or for balance when running. Birds also have pneumatic skeletons (skeletons where bones have substantial air pockets inside them), a feature once interpreted as lightening the skeleton for flight. However, some of the heaviest dinosaurs of all time also had this feature and it is probably an ancient dinosaurian feature. Birds also have a one-way lung system in which inhaled air runs in a “loop” through the body, completely flushing the lungs of old air. Again, this was originally interpreted as an adaptation to the rigors of flying, but is now known to be much more common in the diapsids and not restricted to flighted forms (Schachner et al., 2014; Farmer, 2015a; Farmer, 2015b).
Early birds developed beaks without losing teeth. Until the end of the Mesozoic many lineages of birds had teeth in part of their mouths and had beaks only at the rostral end of the jaws. In the great extinction that ended the Mesozoic all of the toothed birds went extinct, leaving only toothless, beaked forms today. However, the advantages of teeth are such that several modern bird groups have tooth-like serrations in the beaks.
Modern birds also lack bony tails. Early birds would have had long reptilian tails covered in feathers. Modern birds have reduced the bony and fleshy structure of the tail so that most of the tail is composed entirely of feathers. This also means that tail shape is flexible, since birds can flex and fan the feathers to create different tail shapes. This is especially apparent when a flying bird comes in to land, a point at which it often fans the tail to create a wider “brake” in the air.
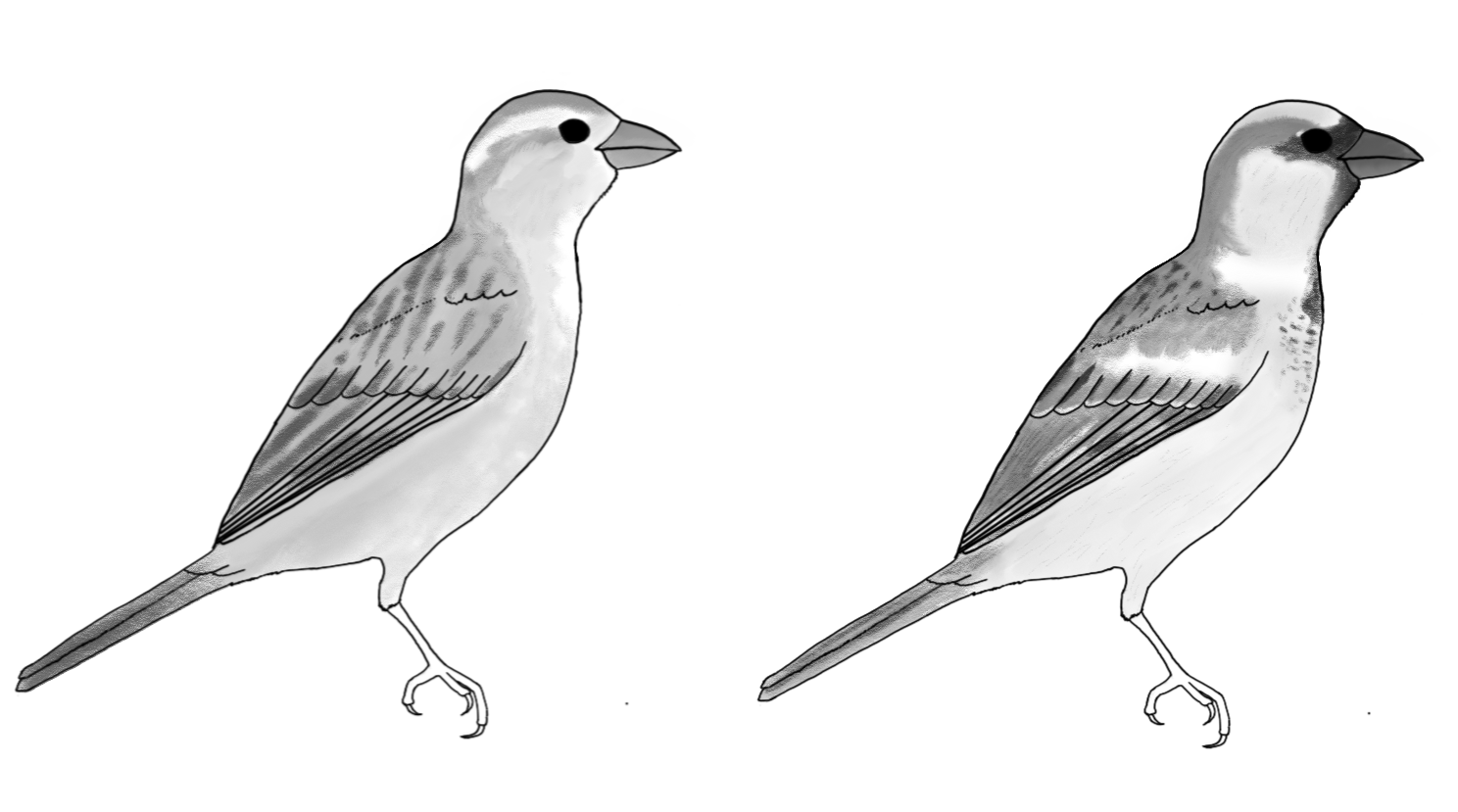
In terms of general morphology, birds are most visibly variable in beak shape (which correlates to diet), wing shape (which varies by flight style), and neck and length. For some birds, such as swifts and hummingbirds, the legs are so reduced as to prevent the bird from walking, and the bird either flies or perches in place. In a sample of six Alpine swifts (Tachymarptis melba) three individuals were found to remain airborne continuously throughout a six-month period (Liechti et al., 2013). In other birds the legs are longer and stronger, indicating that the species spends substantial time on the ground. In many galliform birds (turkeys, quail, grouse, junglefowl) flight is used only for brief periods of time, to escape from predators or move into trees to sleep. In a few species, scattered across the avian phylogeny, the wings are so reduced that flight is impossible and walking or running is the only available mode of locomotion.
Because of the constraints of flight birds tend to be small. The largest birds in the world are all flightless, and it is not until one has passed the ostriches (genus Struthio, 100 kg on average, 220.4 lbs), cassowaries (genus Casuarius), emus (Dromaius novaehollandiae) rheas (genus Rhea), and the world’s largest penguin (the emperor penguin, Aptenodytes forsteri) that one reaches a flying bird. The kori bustard (Ardeotis kori) and great bustard (Otis tarda), both have maximum masses just slightly above 20 kg (44 lbs) and are both considered contenders for the heaviest flying bird, although the lighter wandering albatross (Diomedea exulans) has a much larger wingspan, at 3.65 m (12.0 ft) and stays in flight for much longer periods of time. The smallest bird is generally agreed to be the bee hummingbird at around 2.6 g (0.092 oz).
While flight creates an upper limit to the weight of a bird (specifically, the energy of launch into flight constrains bird body mass) endothermy constrains the lower limit of bird size. As endotherms, birds burn energy for heat, and small animals lose heat much more rapidly than large ones because of the square-cube scaling rule. At some point the energetic costs of heating a small body become impossibly high. Birds tend to have higher body temperatures than mammals.
Birds and mammals have independently evolved four-chambered hearts and complex brains. While the phrase “bird brain” is used derogatorily birds pack neurons into their brains at higher densities than mammals (Olkowicz et al., 2016) and have been shown to perform complex cognitive tasks. For instance, Alex the African gray parrot (Psittacus erithacus) was successfully taught not only to retrieve objects identified only by verbal commands but also to understand abstract class categories and to be able to respond intelligibly to questions using these concepts (Pepperberg, 1983). New Caledonian crows (Corvus moneduloides) have shown the ability to perform metatool tasks, tasks in which a crow must use a tool to get a tool to get a reward. Moreover, most crows grasped this task immediately and performed it without trial and error (Taylor et al., 2007).
Birds are found almost everywhere on earth except deep underwater. Because flight is an efficient way to move long distances birds are able to make use of ephemeral habitats, migrating into an area while conditions are good (during summer or a rainy season) and leaving when conditions get worse. The Arctic tern (Sterna paradisaea), a small seabird, migrates further than any known animal on earth, migrating every year from the Arctic to the oceans around Antarctica, a migration of more than 80,000 km (49,710 miles) (Egevang et al., 2010).
Birds also have a wide variety of diets. Eagles and hawks (Accipitriformes), owls (Strigiformes), and falcons (Falconiformes) are all predators of other terrestrial vertebrates (in larger species). A wide variety of seabirds, herons, loons, and some diving ducks feed on aquatic vertebrates and larger invertebrates. Woodpeckers (Picidae) feed on insects buried inside wood by using a sharp beak and skull equipped with shock-absorbers to hammer into the wood and then use a long tongue that wraps around the base of the bird’s own skull to fish out the insect. Many birds are generalists, eating insects, seeds, and other small, nutritious items. Others specialize on just one of these items. Some birds find food entirely on the wing (swallows and swifts, which feeding on insects while in flight). Grazers and browsers are under-represented in birds, perhaps because the large digestive tracts required to break down low-quality vegetation, but many geese are primarily grazers.
All birds are internally-fertilizing, although the vast majority of birds lack an intromittent organ. In many species reproductive tissues change dramatically in size between breeding and non-breeding season, probably because of the weight limits associated with flight. Courtship in birds is often observed by humans and includes bird songs and colorful displays. Nest-building may be part of courtship, and bird nests range from nothing at all to a few sticks to elaborate structures that are added to year after year. All birds are oviparous and parental care by at least one, and often both, parents is found in all birds except the scrub fowl (Megapodiidae). Newly hatched birds may be precocial (able to move about and feed themselves shortly after hatching, as in chickens) or altricial (unable to move about, feed themselves, and often thermoregulate effectively for a long time after hatching, as in songbirds). Birds that have altricial offspring tend to make nests in protected places, and birds that nest directly on the ground tend to have precocial offspring.
Synapsida (1.0.0)
The story of the synapsids is a story defined by two global disasters. In the Permian the synapsids were a diverse group filling many large-animal niches in terrestrial ecosystems. A new group of synapsids, the therapsids, rose to dominance in the middle of the Permian and the synapsid future looked bright. And then the Permian-Triassic extinction occurred. This extinction, which appears to have been extremely severe and worldwide, decimated the therapsids. In the Triassic the therapsids, now a small portion of their former abundance, were upstaged by a new group of archosaurian diapsids, the dinosaurs. Living in the shadow of the dinosaurs the therapsids continued to evolve, mostly as small, probably nocturnal animals. The Permian therapsids already probably had hair (Bajdek et al., 2016) and in the middle of the Mesozoic the first mammals appeared. While Mesozoic mammals evolved to fill a number of niches they remained small-bodied, kept out of larger-bodied niches by the dinosaurs. And then, 66 million years ago at the end of the Cretaceous, another global disaster occurred. The non-avian dinosaurs, the pterosaurs, the mosasaurs, and the plesiosaurs all went extinct, and all surviving groups of vertebrates lost species, sometimes large percentages of them. However, for the synapsids this created an evolutionary opportunity, and the mammals began evolving to fill the open dinosaurian niches. Today, while the living dinosaurs (birds) have almost twice as many species as the mammals it is the mammals that fill almost all the large-bodied terrestrial niches on earth.
Mammalia (1.0.0)
(Traditionally a Class, approximately 5,852 species, 8.76% of vertebrate species)
The mammals have fewer species than the birds, the squamates, the amphibians, and (of course) the actinopterygians. Despite this, mammals are one of the most morphologically diverse of the traditionally Linnean classes. While the “standard” mammal must be a rodent (rodents are around 40% of all mammal species) the next largest group, the bats, are adapted for flight. The whales are fully aquatic and include the largest animal, by weight, ever to live, the blue whale (Balaenoptera musculus) at 190 tonnes (209 tons) and 33 m (108 ft) (McClain et al., 2015).
Despite mammalian diversity all mammals share a few key traits. All mammals have hair (or fur), thin, keratinous fibers that grow from specialized follicles in the skin. The most common use for hair is insulation, as all mammals are endotherms. However, hair is also used for sensing (whiskers, properly a sub-type of vibrissae), defense (spines and quills), and display. While some mammals, such as whales, hippos, rhinos, and elephants, are largely hairless all mammals do have at least a small number of functioning hair follicles. In some “hairless” mammals the insulative hairs are lost but sensory hairs are still present, as on whales (Drake et al., 2015).
All mammals also give milk to their offspring. Milk allows a mother mammal to transfer energy and nutrients from herself to the offspring after they are born but also necessitates parental care, which is therefore universal in mammals. Milk varies widely in percentages of fat, protein, and carbohydrates. For instance, cow’s milk is 3-4% fat while the milk of a blue whale, whose calf can grow at 113 kg (250 lbs) a day, can have up to 49% fat (Gregory et al., 1955). Mammary glands are thought to be derived from sweat glands, another mammalian characteristic.
Mammals also have a distinctive ear, with three ossicles (malleus, incus, and stapes) that connect the eardrum to the inner ear. This is paired with a unique jaw joint. These skeletal features are important in determining whether a fossil animal is a mammal.
Mammals also have distinctive teeth (amongst living species – the features described here are not all unique to mammals when extinct synapsids are also considered). First, most mammals have several types of teeth in their mouths. (Several groups of mammals have no teeth at all, and in a few groups, such as the toothed whales, all teeth are of one type.) The basic tooth types are incisors (front teeth, used for slicing), canine teeth (beside the incisors, stabbing teeth, used for fighting or grabbing/killing struggling prey), premolars (between the canines and molars, two cusps, used for chewing), and molars (more than two cusps, used for more extensive chewing/grinding). Variations in this pattern are common. For instance, many groups of mammals do not have canines at all. In some cases teeth are modified well beyond these basic types. Elephant and narwhal tusks are teeth.
Second, while in many toothed species tooth replacement is a constant process (individual teeth fall out and are replaced without a schedule) in mammals tooth replacement is a simultaneous process. In most mammals all teeth are replaced once, when the “baby teeth” are replaced with adult teeth, the process seen in humans. In a few mammals teeth are never replaced at all. A few mammals have a continuous tooth replacement process (elephants, for instance) but this appears to have evolved from ancestors who had a more normal mammalian pattern.
The diversity in mammalian teeth has allowed mammals to adopt many different diets. A large mammalian clade called Carnivora includes all the large, terrestrial mammalian predators, and is composed of the dogs, bears, cats, seals, weasel family, raccoon family, hyenas, mongooses, and civets. Even in this “carnivorous” group there are many omnivores and a few herbivores. Other clade are strict herbivores, including some very efficient grazers and browsers that can process vegetation so well that they can survive on only grasses and other low-quality plant foods. In the baleen whales teeth are absent and filter plates, called baleen, are used in filter feeding. Rodents, the most diverse group, have evolved ever-growing incisors that allow them to gnaw on hard food (or to use their teeth to carve out shelters) without wearing their teeth down and starving.
Mammals are placed into three large groups based on their reproductive modes. These reproductive modes correlate to large phylogenetic splits, meaning that while we may separate these groups based on reproductive mode they are also monophyletic groups that may share many other features.
Monotremes
Monotremes are composed of only five living species, the platypus (Ornithorhynchus anatinus) and the four echidnas (Tachyglossidae), all of which are found in Australasia. Monotremes are oviparous, laying eggs that hatch into under-developed offspring that then require significant parental care before they can move on their own. Monotremes are also notable for having a cloaca, a single urogenital opening (“monotreme” is a name that refers to this one opening), unlike other mammals. Monotremes are the sister clade to all other mammals.
Marsupials
Marsupials are a much larger group than monotremes with around 327 species, although still a small fraction of living mammals. Modern marsupials are restricted to the Americas and Australasia. The Australian marsupials are notable for having evolved to fill a huge range of niches, with marsupials that are large predators (the thylacine, Thylacinus cynocephalus), large herbivores (various kangaroos), medium-sized predators (the quolls and the Tasmanian devil), ant and termite eaters (the numbat, Myrmecobius fasciatus), mole-equivalents (marsupial moles, genus Notoryctes), burrowing herbivores (the wombats), and numerous others. The American marsupials are largely restricted to South America, with the number of species dropping sharply as one moves into Central America and further north. The American marsupials (most of which have common names involving the word “opossum”) are largely generalist omnivores, and most can climb to some degree, although in both omnivory and climbing ability there are wide variations, with some mostly-herbivorous species, some mostly-carnivorous species, some largely arboreal species, and some largely terrestrial species. Despite this pattern, the American marsupials are the older clade. Australian marsupials are all descended from some members of some species within the American marsupial clade that made their way across Antarctica (which was, at that time, further north, temperate in climate, and joined to both South America and Australia) and into Australia.
Marsupials are viviparous, but the young are born after a much shorter time than in the more familiar placental mammals. These underdeveloped young then move into their mother’s pouch, latch onto a teat, and remain there until they have developed sufficiently to leave the pouch. This allows a female marsupial to have more broods of offspring in quick succession than her placental counterparts (she can have a brood in utero and a brood in the pouch at developmental stages where a placental mammal would still have those offspring in utero) but the early demands of climbing to the pouch and feeding from the mother’s teat have been proposed as fundamental constraints on marsupials that have prevented them from diversifying further. However, this issue is still hotly debated, and it is also possible that placental dominance is largely a random artifact of events in earth’s history (Sánchez-Villagra, 2013).
Placentals/Eutherians
The group known as placentals or eutherians (both names refer to the same group of living mammals, although they include different extinct species) are the majority of mammals. Placentals are viviparous and do not have any distinct post-birth stage as in marsupials. Some placentals are relatively helpless at birth (dogs, humans) while others are able to see, run, and move with adults within hours of birth (horses, many antelope).
References
Aalbers, S. A., Bernal, D., & Sepulveda, C. A. (2010). The functional role of the caudal fin in the feeding ecology of the common thresher shark Alopias vulpinus. Journal of Fish Biology, 76(7), 1863–1868. https://doi.org/10.1111/j.1095-8649.2010.02616.x
Allen, G. R., Midgley, S. H., & Allen, M. (2002). Field guide to the freshwater fishes of Australia. Western Australian Museum.
Anderson Philip S.L, & Westneat Mark W. (2007). Feeding mechanics and bite force modelling of the skull of Dunkleosteus terrelli, an ancient apex predator. Biology Letters, 3(1), 77–80. https://doi.org/10.1098/rsbl.2006.0569
Arthur, K. E., Boyle, M. C., & Limpus, C. J. (2008). Ontogenetic changes in diet and habitat use in green sea turtle (Chelonia mydas) life history. Marine Ecology Progress Series, 362, 303–311. https://doi.org/10.3354/meps07440
Babiker, M. M. (1979). Respiratory behaviour, oxygen consumption and relative dependence on aerial respiration in the African lungfish (Protopterus annectens, owen) and an air-breathing teleost (Clarias lazera, C.). Hydrobiologia, 65(2), 177–187. https://doi.org/10.1007/BF00017423
Bajdek, P., Qvarnström, M., Owocki, K., Sulej, T., Sennikov, A. G., Golubev, V. K., & Niedźwiedzki, G. (2016). Microbiota and food residues including possible evidence of pre-mammalian hair in Upper Permian coprolites from Russia. Lethaia, 49(4), 455–477. https://doi.org/10.1111/let.12156
Bardack, D. (1991). First Fossil Hagfish (Myxinoidea): A Record from the Pennsylvanian of Illinois. Science, 254(5032), 701–703. https://doi.org/10.1126/science.254.5032.701
Bhattarai, S. (2015). Notes on Mugger Crocodile Crocodylus palustris (Lesson, 1831) hunting on Axis axis in Bardia National Park, Nepal. Hyla : Herpetological Bulletin, 2015(2), 41–44.
Binion, G. R., Daughtery, D. J., & Bodine, K. A. (2015). Population Dynamics of Alligator Gar in Choke Canyon Reservoir, Texas: Implications for Management. Journal of the Association of Southeastern Fish and Wildlife Agencies, 2, 57–63.
Blair, J. E., & Hedges, S. B. (2005). Molecular phylogeny and divergence times of deuterostome animals. Molecular Biology and Evolution, 22(11), 2275–2284. https://doi.org/10.1093/molbev/msi225
Blake, S., Wikelski, M., Cabrera, F., Guezou, A., Silva, M., Sadeghayobi, E., … Jaramillo, P. (2012). Seed dispersal by Galápagos tortoises. Journal of Biogeography, 39(11), 1961–1972. https://doi.org/10.1111/j.1365-2699.2011.02672.x
Boback, S., Snow, R., Hsu, T., Peurach, S., Dove, C., & Reed, R. (2016). Supersize Me: Remains of Three White-Tailed Deer (Odocoileus virginianus) in an Invasive Burmese Python (Python molurus bivittatus) in Florida. BioInvasions Records, 5(4), 197–203. https://doi.org/10.3391/bir.2016.5.4.02
Bone, Q., & Moore, R. H. . (2008). Biology of Fishes (Third Edit). New York, New York: Taylor & Francis.
Booth, W., Smith, C. F., Eskridge, P. H., Hoss, S. K., Mendelson, J. R., & Schuett, G. W. (2012). Facultative parthenogenesis discovered in wild vertebrates. Biology Letters, 8(6), 983–985. https://doi.org/10.1098/rsbl.2012.0666
Branch, W. R. (1976). Two Exceptional Food Records for the African Bullfrog, Pyxicephalus adspersus (Amphibia, Anura, Ranidae). Journal of Herpetology, 10(3), 266–268. https://doi.org/10.2307/1562997
Britton, A. R. C., Whitaker, R., & Whitaker, N. (2012). Here be a Dragon: Exceptional Size in a Saltwater Crocodile (Crocodylus porosus) from the Philippines. Herpetological Reviews, 43(4), 541–546.
Buckley, D., Alcobendas, M., García‐París, M., & Wake, M. H. (2007). Heterochrony, cannibalism, and the evolution of viviparity in Salamandra salamandra. Evolution & Development, 9(1), 105–115. https://doi.org/10.1111/j.1525-142X.2006.00141.x
Bull, J. J., & Vogt, R. C. (1979). Temperature-dependent sex determination in turtles. Science, 206(4423), 1186–1188. https://doi.org/10.1126/science.505003
Bullock, T. H., Moore, J. K., & Fields, R. D. (1984). Evolution of myelin sheaths: Both lamprey and hagfish lack myelin. Neuroscience Letters, 48(2), 145–148. https://doi.org/10.1016/0304-3940(84)90010-7
Burton, T. M., & Likens, G. E. (1975). Salamander Populations and Biomass in the Hubbard Brook Experimental Forest, New Hampshire. Copeia, 1975(3), 541–546. https://doi.org/10.2307/1443655
Buscalioni, A. D., Ortega, F., & Vasse, D. (1997). New crocodiles (Eusuchia: Alligatoroidea) from the Upper Cretaceous of southern Europe. Comptes Rendus de l’Académie Des Sciences – Series IIA – Earth and Planetary Science, 325(7), 525–530. https://doi.org/10.1016/S1251-8050(97)89872-2
Cardwell, J. R., & Liley, N. R. (1991). Hormonal control of sex and color change in the stoplight parrotfish, Sparisoma viride. General and Comparative Endocrinology, 81(1), 7–20. https://doi.org/10.1016/0016-6480(91)90120-U
Chapman Demian D., Wintner Sabine P., Abercrombie Debra L., Ashe Jimiane, Bernard Andrea M., Shivji Mahmood S., & Feldheim Kevin A. (2013). The behavioural and genetic mating system of the sand tiger shark, Carcharias taurus, an intrauterine cannibal. Biology Letters, 9(3), 20130003. https://doi.org/10.1098/rsbl.2013.0003
Chavez, G., Venegas, P. J., & Lescano, A. (2011). Two new records in the diet of Ceratophrys cornuta Linneaus, 1758 (Anura:Ceratophrydae). Herpetology Notes, 4, 285–286.
Clark, A. J., & Summers, A. P. (2007). Morphology and kinematics of feeding in hagfish: possible functional advantages of jaws. Journal of Experimental Biology, 210(22), 3897–3909. https://doi.org/10.1242/jeb.006940
Colli, G. R., & Zamboni, D. S. (1999). Ecology of the Worm-Lizard Amphisbaena alba in the Cerrado of Central Brazil. Copeia, 1999(3), 733–742. https://doi.org/10.2307/1447606
Córdoba, E. A., León, Á. V. J., Bonilla-Castillo, C. A., Petrere Junior, M., Peláez, M., Duponchelle, F., … Duponchelle, F. (2013). Breeding, growth and exploitation of Brachyplatystoma rousseauxii Castelnau, 1855 in the Caqueta River, Colombia. Neotropical Ichthyology, 11(3), 637–647. https://doi.org/10.1590/S1679-62252013000300017
Crawford, N. G., Faircloth, B. C., McCormack, J. E., Brumfield, R. T., Winker, K., & Glenn, T. C. (2012). More than 1000 ultraconserved elements provide evidence that turtles are the sister group of archosaurs. Biology Letters, 8(5), 783–786. https://doi.org/10.1098/rsbl.2012.0331
Cree, A., Cockrem, J. F., & Guillette, L. J. (1992). Reproductive cycles of male and female tuatara (Sphenodon punctatus) on Stephens Island, New Zealand. Journal of Zoology, 226(2), 199–217. https://doi.org/10.1111/j.1469-7998.1992.tb03834.x
CROOK, I. G. (1973). THE TUATARA, SPHENODON PUNCTATUS GRAY, ON ISLANDS WITH AND WITHOUT POPULATIONS OF THE POLYNESIAN RAT, RATTUS EXULANS (PEALE). Proceedings (New Zealand Ecological Society), 20, 115–120.
Dean, M. N., & Summers, A. P. (2006). Mineralized cartilage in the skeleton of chondrichthyan fishes. Zoology, 109(2), 164–168. https://doi.org/10.1016/j.zool.2006.03.002
Delany, M. F., & Abercrombie, C. L. (1986). American Alligator Food Habits in Northcentral Florida. The Journal of Wildlife Management, 50(2), 348–353. https://doi.org/10.2307/3801926
Dinets, V., Brueggen, J. C., & Brueggen, J. D. (2015). Crocodilians use tools for hunting. Ethology Ecology & Evolution, 27(1), 74–78. https://doi.org/10.1080/03949370.2013.858276
Doherty, A. H., Ghalambor, C. K., & Donahue, S. W. (2015). Evolutionary Physiology of Bone: Bone Metabolism in Changing Environments. Physiology, 30(1), 17–29. https://doi.org/10.1152/physiol.00022.2014
Donley, J. M., Sepulveda, C. A., Konstantinidis, P., Gemballa, S., & Shadwick, R. E. (2004). Convergent evolution in mechanical design of lamnid sharks and tunas. Nature, 429(6987), 61–65. https://doi.org/10.1038/nature02435
dos Reis, M., Thawornwattana, Y., Angelis, K., Telford, M. J., Donoghue, P. C. J., & Yang, Z. (2015). Uncertainty in the Timing of Origin of Animals and the Limits of Precision in Molecular Timescales. Current Biology: CB, 25(22), 2939–2950. https://doi.org/10.1016/j.cub.2015.09.066
Drake, S. E., Crish, S. D., George, J. C., Stimmelmayr, R., & Thewissen, J. G. M. (2015). Sensory Hairs in the Bowhead Whale, Balaena mysticetus (Cetacea, Mammalia). The Anatomical Record, 298(7), 1327–1335. https://doi.org/10.1002/ar.23163
Egevang, C., Stenhouse, I. J., Phillips, R. A., Petersen, A., Fox, J. W., & Silk, J. R. D. (2010). Tracking of Arctic terns Sterna paradisaea reveals longest animal migration. Proceedings of the National Academy of Sciences, 107(5), 2078–2081. https://doi.org/10.1073/pnas.0909493107
Farmer, C. G. (2015a). Similarity of Crocodilian and Avian Lungs Indicates Unidirectional Flow Is Ancestral for Archosaurs. Integrative and Comparative Biology, 55(6), 962–971. https://doi.org/10.1093/icb/icv078
Farmer, C. G. (2015b). The Evolution of Unidirectional Pulmonary Airflow. Physiology, 30(4), 260–272. https://doi.org/10.1152/physiol.00056.2014
Fernholm, B., & Holmberg, K. (1975). The eyes in three genera of hagfish (Eptatretus, paramyxine andMyxine)—A case of degenerative evolution. Vision Research, 15(2), 253-IN4. https://doi.org/10.1016/0042-6989(75)90215-1
Fredriksson, G. M. (2005). PREDATION ON SUN BEARS BY RETICULATED PYTHON IN EAST KALIMANTAN, INDONESIAN BORNEO. The Raffles Bulletin of Zoology, 53(1), 165–168.
Fricke, H., Hissmann, K., Schauer, J., Reinicke, O., Kasang, L., & Plante, R. (1991). Habitat and population size of the coelacanth Latimeria chalumnae at Grand Comoro. Environmental Biology of Fishes, 32(1), 287–300. https://doi.org/10.1007/BF00007462
Friedman, M., & Coates, M. I. (2006). A newly recognized fossil coelacanth highlights the early morphological diversification of the clade. Proceedings of the Royal Society B: Biological Sciences, 273(1583), 245–250. https://doi.org/10.1098/rspb.2005.3316
Fry, B. G., Wroe, S., Teeuwisse, W., Osch, M. J. P. van, Moreno, K., Ingle, J., … Norman, J. A. (2009). A central role for venom in predation by Varanus komodoensis (Komodo Dragon) and the extinct giant Varanus (Megalania) priscus. Proceedings of the National Academy of Sciences, pnas.0810883106. https://doi.org/10.1073/pnas.0810883106
Gaitonde, N., & Giri, V. (2014). Primitive breeding in an ancient Indian frog genus Indirana. Current Science, 107(1), 109–112.
Gerringer, M. E., Linley, T. D., Jamieson, A. J., Goetze, E., & Drazen, J. C. (2017). Pseudoliparis swirei sp. nov.: A newly-discovered hadal snailfish (Scorpaeniformes: Liparidae) from the Mariana Trench. Zootaxa, 4358(1), 161–177. https://doi.org/10.11646/zootaxa.4358.1.7
Gess, R. W., Coates, M. I., & Rubidge, B. S. (2006). A lamprey from the Devonian period of South Africa. Nature, 443(7114), 981–984. https://doi.org/10.1038/nature05150
Giacomo, E. D., & Perier, M. R. (1996). Feeding habits of cockfish, Callorhinchus callorhynchus (Holocephali: Callorhynchidae), in Patagonian waters (Argentina). Marine and Freshwater Research, 47(6), 801–808. https://doi.org/10.1071/mf9960801
Gibbons, J. W., Semlitsch, R. D., Greene, J. L., & Schubauer, J. P. (1981). Variation in Age and Size at Maturity of the Slider Turtle (Pseudemys scripta). The American Naturalist, 117(5), 841–845. https://doi.org/10.1086/283774
Glaw, F., Köhler, J., Townsend, T. M., & Vences, M. (2012). Rivaling the World’s Smallest Reptiles: Discovery of Miniaturized and Microendemic New Species of Leaf Chameleons (Brookesia) from Northern Madagascar. PLOS ONE, 7(2), e31314. https://doi.org/10.1371/journal.pone.0031314
Godwin, J. R., & Thomas, P. (1993). Sex Change and Steroid Profiles in the Protandrous Anemonefish Amphiprion melanopus (Pomacentridae, Teleostei). General and Comparative Endocrinology, 91(2), 144–157. https://doi.org/10.1006/gcen.1993.1114
Goicoechea, O., Garrido, O., & Jorquera, B. (1986). Evidence for a Trophic Paternal-Larval Relationship in the Frog Rhinoderma darwinii. Journal of Herpetology, 20(2), 168–178. https://doi.org/10.2307/1563941
Goldman, K. J. (1997). Regulation of body temperature in the white shark, Carcharodon carcharias. Journal of Comparative Physiology B, 167(6), 423–429. https://doi.org/10.1007/s003600050092
Gower, D. J., Giri, V., Dharne, M. S., & Shouche, Y. S. (2008). Frequency of independent origins of viviparity among caecilians (Gymnophiona): evidence from the first ‘live-bearing’ Asian amphibian. Journal of Evolutionary Biology, 21(5), 1220–1226. https://doi.org/10.1111/j.1420-9101.2008.01577.x
Graham, R. T., & Roberts, C. M. (2007). Assessing the size, growth rate and structure of a seasonal population of whale sharks (Rhincodon typus Smith 1828) using conventional tagging and photo identification. Fisheries Research, 84(1), 71–80. https://doi.org/10.1016/j.fishres.2006.11.026
Greene, H. W., Rodríguez, J. J. S., & Powell, B. J. (2006). PARENTAL BEHAVIOR IN ANGUID LIZARDS. South American Journal of Herpetology, 1(1), 9–20. https://doi.org/10.2994/1808-9798(2006)1[9:PBIAL]2.0.CO;2
Gregory, M. E., Kon, S. K., Rowland, S. J., & Thompson, S. Y. (1955). 578. The composition of the milk of the blue whale. Journal of Dairy Research, 22(1), 108–112. https://doi.org/10.1017/S0022029900007615
Halstead, B. W., & Bunker, N. C. (1952). The Venom Apparatus of the Ratfish, Hydrolagus colliei. Copeia, 1952(3), 128–138. https://doi.org/10.2307/1439692
Harel, I., Benayoun, B. A., Machado, B., Singh, P. P., Hu, C.-K., Pech, M. F., … Brunet, A. (2015). A platform for rapid exploration of aging and diseases in a naturally short-lived vertebrate. Cell, 160(5), 1013–1026. https://doi.org/10.1016/j.cell.2015.01.038
Hart, D. R. (1983). Dietary and Habitat Shift with Size of Red-Eared Turtles (Pseudemys scripta) in a Southern Louisiana Population. Herpetologica, 39(3), 285–290.
Heimberg, A. M., Cowper-Sal·lari, R., Sémon, M., Donoghue, P. C. J., & Peterson, K. J. (2010). microRNAs reveal the interrelationships of hagfish, lampreys, and gnathostomes and the nature of the ancestral vertebrate. Proceedings of the National Academy of Sciences, 107(45), 19379–19383. https://doi.org/10.1073/pnas.1010350107
Hilliard, R. W., Potter, I. C., & Macey, D. J. (1985). The Dentition and Feeding Mechanism in Adults of the Southern Hemisphere Lamprey Geotria australis Gray. Acta Zoologica, 66(3), 159–170. https://doi.org/10.1111/j.1463-6395.1985.tb00834.x
Huber Daniel R, Dean Mason N, & Summers Adam P. (2008). Hard prey, soft jaws and the ontogeny of feeding mechanics in the spotted ratfish Hydrolagus colliei. Journal of The Royal Society Interface, 5(25), 941–953. https://doi.org/10.1098/rsif.2007.1325
Inoue, J. G., Miya, M., Venkatesh, B., & Nishida, M. (2005). The mitochondrial genome of Indonesian coelacanth Latimeria menadoensis (Sarcopterygii: Coelacanthiformes) and divergence time estimation between the two coelacanths. Gene, 349, 227–235. https://doi.org/10.1016/j.gene.2005.01.008
James, M. C., & Mrosovsky, N. (2004). Body temperatures of leatherback turtles (Dermochelys coriacea) in temperate waters off Nova Scotia, Canada. Canadian Journal of Zoology, 82(8), 1302–1306. https://doi.org/10.1139/z04-110
Jessop, T. S., Madsen, T., Sumner, J., Rudiharto, H., Phillips, J. A., & Ciofi, C. (2006). Maximum body size among insular Komodo dragon populations covaries with large prey density. Oikos, 112(2), 422–429. https://doi.org/10.1111/j.0030-1299.2006.14371.x
Kearney, M., Fujita, M. K., & Ridenour, J. (2009). Lost Sex in the Reptiles: Constraints and Correlations. In I. Schön, K. Martens, & P. Dijk (Eds.), Lost Sex: The Evolutionary Biology of Parthenogenesis (pp. 447–474). Dordrecht: Springer Netherlands. https://doi.org/10.1007/978-90-481-2770-2_21
Kottelat, M. (2001). Fishes of Laos. WHT Publications Ltd., Colombo 5, Sri Lanka.
Kupfer, A., Müller, H., Antoniazzi, M. M., Jared, C., Greven, H., Nussbaum, R. A., & Wilkinson, M. (2006). Parental investment by skin feeding in a caecilian amphibian. Nature, 440(7086), 926–929. https://doi.org/10.1038/nature04403
Kuraku, S., & Kuratani, S. (2006). Time Scale for Cyclostome Evolution Inferred with a Phylogenetic Diagnosis of Hagfish and Lamprey cDNA Sequences. Zoological Science, 23(12), 1053–1064. https://doi.org/10.2108/zsj.23.1053
Kuratani, S. (2004). Evolution of the vertebrate jaw: comparative embryology and molecular developmental biology reveal the factors behind evolutionary novelty. Journal of Anatomy, 205(5), 335–347. https://doi.org/10.1111/j.0021-8782.2004.00345.x
Lang, J. W., & Andrews, H. V. (1994). Temperature-dependent sex determination in crocodilians. Journal of Experimental Zoology, 270(1), 28–44. https://doi.org/10.1002/jez.1402700105
Law, S. J., De Kort, S. R., & van Weerd, M. (2016). Morphology, Activity Area, and Movement Patterns of the frugivorous monitor lizard Varanus bitatawa. Herpetological Conservation and Biology, 11, 467–475.
Liechti, F., Witvliet, W., Weber, R., & Bächler, E. (2013). First evidence of a 200-day non-stop flight in a bird. Nature Communications, 4, 2554. https://doi.org/10.1038/ncomms3554
Lloyd, G. T., Wang, S. C., & Brusatte, S. L. (2012). Identifying Heterogeneity in Rates of Morphological Evolution: Discrete Character Change in the Evolution of Lungfish (sarcopterygii; Dipnoi). Evolution, 66(2), 330–348. https://doi.org/10.1111/j.1558-5646.2011.01460.x
Longo, S., Riccio, M., & McCune, A. R. (2013). Homology of lungs and gas bladders: Insights from arterial vasculature. Journal of Morphology, 274(6), 687–703. https://doi.org/10.1002/jmor.20128
Lourdais, O., Hoffman, T. C. M., & DeNardo, D. F. (2007). Maternal brooding in the children’s python (Antaresia childreni) promotes egg water balance. Journal of Comparative Physiology B, 177(5), 569–577. https://doi.org/10.1007/s00360-007-0155-6
Lund, R. (1980). Viviparity and Intrauterine Feeding in a New Holocephalan Fish from the Lower Carboniferous of Montana. Science, 209(4457), 697–699. https://doi.org/10.1126/science.209.4457.697
Magnusson, W. E., da Silva, E. V., & Lima, A. P. (1987). Diets of Amazonian Crocodilians. Journal of Herpetology, 21(2), 85–95. https://doi.org/10.2307/1564468
Mallatt, J. (1981). The suspension feeding mechanism of the larval lamprey Petromyzon marinus. Journal of Zoology, 194(1), 103–142. https://doi.org/10.1111/j.1469-7998.1981.tb04581.x
Martín, J., Ortega, J., López, P., Pérez‐Cembranos, A., & Pérez‐Mellado, V. (2013). Fossorial life does not constrain diet selection in the amphisbaenian Trogonophis wiegmanni. Journal of Zoology, 291(3), 226–233. https://doi.org/10.1111/jzo.12064
McClain, C. R., Balk, M. A., Benfield, M. C., Branch, T. A., Chen, C., Cosgrove, J., … Thaler, A. D. (2015). Sizing ocean giants: patterns of intraspecific size variation in marine megafauna. PeerJ, 3, e715. https://doi.org/10.7717/peerj.715
Measey, J. G., & Herrel, A. (2006). Rotational feeding in caecilians: putting a spin on the evolution of cranial design. Biology Letters, 2(4), 485–487. https://doi.org/10.1098/rsbl.2006.0516
Miller, R. F., Cloutier, R., & Turner, S. (2003). The oldest articulated chondrichthyan from the Early Devonian period. Nature, 425(6957), 501–504. https://doi.org/10.1038/nature02001
Moldowan, P. D., Keevil, M. G., Kell, S., Brooks, R. J., & Litzgus, J. D. (2015). Anti-predator defenses of Brown Bullheads ( Ameiurus nebulosus ) and interactions with Snapping Turtles ( Chelydra serpentina ). The Canadian Field-Naturalist, 129(2), 189–193. https://doi.org/10.22621/cfn.v129i2.1701
Muedeking, M. H., & Heyer, W. R. (1976). Descriptions of Eggs and Reproductive Patterns of Leptodactylus pentadactylus (Amphibia: Leptodactylidae). Herpetologica, 32(2), 137–139.
Neira, L. E. F. D., & Johnson, M. K. (1985). Diets of Giant Tortoises and Feral Burros on Volcan Alcedo, Galapagos. The Journal of Wildlife Management, 49(1), 165–169. https://doi.org/10.2307/3801865
Nomenclature, I. C. on Z. (1941). Opinion 114 Under suspension Simia, Simia satyrus and Pithecus are suppressed. Retrieved from https://core.ac.uk/display/33929825
Oftedal, O. T. (2000). Use of maternal reserves as a lactation strategy in large mammals. Proceedings of the Nutrition Society, 59(1), 99–106. https://doi.org/10.1017/S0029665100000124
Olkowicz, S., Kocourek, M., Lučan, R. K., Porteš, M., Fitch, W. T., Herculano-Houzel, S., & Němec, P. (2016). Birds have primate-like numbers of neurons in the forebrain. Proceedings of the National Academy of Sciences, 201517131. https://doi.org/10.1073/pnas.1517131113
Patchell, F. C., & Shine, R. (1986). Food Habits and Reproductive Biology of the Australian Legless Lizards (Pygopodidae). Copeia, 1986(1), 30–39. https://doi.org/10.2307/1444884
Penning, D. A., Dartez, S. F., & Moon, B. R. (2015). The big squeeze: scaling of constriction pressure in two of the world’s largest snakes, Python reticulatus and Python molurus bivittatus. Journal of Experimental Biology, 218(21), 3364–3367. https://doi.org/10.1242/jeb.127449
Pepperberg, I. M. (1983). Cognition in the African Grey parrot: Preliminary evidence for auditory/vocal comprehension of the class concept. Animal Learning & Behavior, 11(2), 179–185. https://doi.org/10.3758/BF03199646
Perry, S. F., Wilson, R. J. A., Straus, C., Harris, M. B., & Remmers, J. E. (2001). Which came first, the lung or the breath? Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 129(1), 37–47. https://doi.org/10.1016/S1095-6433(01)00304-X
Pfennig, D. W. (1992). Polyphenism in Spadefoot Toad Tadpoles as a Locally Adjusted Evolutionarily Stable Strategy. Evolution, 46(5), 1408–1420. https://doi.org/10.1111/j.1558-5646.1992.tb01133.x
Platt, S. G., Elsey, R. M., Liu, H., Rainwater, T. R., Nifong, J. C., Rosenblatt, A. E., … Mazzotti, F. J. (2013). Frugivory and seed dispersal by crocodilians: an overlooked form of saurochory? Journal of Zoology, 291(2), 87–99. https://doi.org/10.1111/jzo.12052
Preez, J. J. du, Jarvis, M. J. F., Capatos, D., & Kock, J. de. (1992). A note on growth curves for the ostrich (Struthio camelus). Animal Science, 54(1), 150–152. https://doi.org/10.1017/S0003356100020687
Punzo, F. (1975). Studies on the Feeding Behavior, Diet, Nesting Habits and Temperature Relationships of Chelydra serpentina osceola (Chelonia: Chelydridae). Journal of Herpetology, 9(2), 207–210. https://doi.org/10.2307/1563038
Roach, J. (2003). World’s heaviest bony fish discovered? National Geographic News, (13 May 2003).
Roberts, T. R. (2012). Systematics, biology, and distribution of the species of the oceanic oarfish genus Regalecus (Teleostei, Lampridiformes, Regalecidae) (Vol. 202). Paris: Mémoires du Muséum national d’histoire naturelle.
Salisbury, S. W., Molnar, R. E., Frey, E., & Willis, P. M. A. (2006). The origin of modern crocodyliforms: new evidence from the Cretaceous of Australia. Proceedings of the Royal Society of London B: Biological Sciences, 273(1600), 2439–2448. https://doi.org/10.1098/rspb.2006.3613
Sanaiotti, T., Magnusson, W., & Campos, Z. (2010). Maximum size of dwarf caiman, Paleosuchus palpebrosus (Cuvier, 1807), in the Amazon and habitats surrounding the Pantanal, Brazil. Amphibia-Reptilia, 31(3), 439–442. https://doi.org/10.1163/156853810791769392
Sánchez-Villagra, M. R. (2013). Why are There Fewer Marsupials than Placentals? On the Relevance of Geography and Physiology to Evolutionary Patterns of Mammalian Diversity and Disparity. Journal of Mammalian Evolution, 20(4), 279–290. https://doi.org/10.1007/s10914-012-9220-3
Schachner, E. R., Cieri, R. L., Butler, J. P., & Farmer, C. G. (2014). Unidirectional pulmonary airflow patterns in the savannah monitor lizard. Nature, 506(7488), 367–370. https://doi.org/10.1038/nature12871
Scott, R., Marsh, R., & Hays, G. C. (2012). Life in the really slow lane: loggerhead sea turtles mature late relative to other reptiles. Functional Ecology, 26(1), 227–235. https://doi.org/10.1111/j.1365-2435.2011.01915.x
Shelton, R. G. J. (1978). On the Feeding of the Hagfish Myxine Glutinosa in the North Sea. Journal of the Marine Biological Association of the United Kingdom, 58(1), 81–86. https://doi.org/10.1017/S0025315400024413
Smith, C. L., Rand, C. S., Schaeffer, B., & Atz, J. W. (1975). Latimeria, the Living Coelacanth, Is Ovoviviparous. Science, 190(4219), 1105 LP – 1106.
Stone, J., & Lohman, K. (2006). Observations on nest characteristics, spawning habitat, and spawning behavior of pacific and western brook lamprey in a washington stream. Northwestern Naturalist, 87(3), 225–232. https://doi.org/10.1898/1051-1733(2006)87[225:OONCSH]2.0.CO;2
Summers, A. P. (2000). Stiffening the stingray skeleton — an investigation of durophagy in Myliobatid stingrays (Chondrichthyes, Batoidea, Myliobatidae). Journal of Morphology, 243(2), 113–126. https://doi.org/10.1002/(SICI)1097-4687(200002)243:2<113::AID-JMOR1>3.0.CO;2-A
Taylor, A. H., Hunt, G. R., Holzhaider, J. C., & Gray, R. D. (2007). Spontaneous Metatool Use by New Caledonian Crows. Current Biology, 17(17), 1504–1507. https://doi.org/10.1016/j.cub.2007.07.057
Tortonese, E. (1986). Molidae. In P. J. P. Whitehead, M.-L. Bauchot, J.-C. Hureau, J. Nielsen, & E. Tortonese (Eds.), Fishes of the North-eastern Atlantic and the Mediterranean (Vol. 3, pp. 1348–1350). UNESCO, Paris.
Townsend, T. M., Larson, A., Louis, E., & Macey, J. R. (2004). Molecular Phylogenetics of Squamata: The Position of Snakes, Amphisbaenians, and Dibamids, and the Root of the Squamate Tree. Systematic Biology, 53(5), 735–757. https://doi.org/10.1080/10635150490522340
Troyer, K. (1984). Diet selection and digestion in Iguana iguana: the importance of age and nutrient requirements. Oecologia, 61(2), 201–207. https://doi.org/10.1007/BF00396761
Tschopp, E., Mateus, O., & Benson, R. B. J. (2015). A specimen-level phylogenetic analysis and taxonomic revision of Diplodocidae (Dinosauria, Sauropoda). PeerJ, 3, e857. https://doi.org/10.7717/peerj.857
Twohey, M. B., Sorensen, P. W., & Li, W. (2003). Possible Applications of Pheromones in an Integrated Sea Lamprey Management Program. Journal of Great Lakes Research, 29, 794–800. https://doi.org/10.1016/S0380-1330(03)70532-2
Underwood, C. J. (2006). Diversification of the Neoselachii (Chondrichthyes) during the Jurassic and Cretaceous. Paleobiology, 32(2), 215–235. https://doi.org/10.1666/04069.1
Verdade, V. K., Schiesari, L. C., & Bertoluci, J. A. (2000). Diet of Juvenile Aquatic Caecilians, Typhlonectes compressicauda. Journal of Herpetology, 34(2), 291–293. https://doi.org/10.2307/1565428
Vidal, N., & Hedges, S. B. (2009). The molecular evolutionary tree of lizards, snakes, and amphisbaenians. Comptes Rendus Biologies, 332(2), 129–139. https://doi.org/10.1016/j.crvi.2008.07.010
Vrtílek, M., Žák, J., Pšenička, M., & Reichard, M. (2018). Extremely rapid maturation of a wild African annual fish. Current Biology, 28(15), R822–R824. https://doi.org/10.1016/j.cub.2018.06.031
Wallace, K. M., & Leslie, A. J. (2008). Diet of the Nile Crocodile (Crocodylus niloticus) in the Okavango Delta, Botswana. Journal of Herpetology, 42(2), 361–369. https://doi.org/10.1670/07-1071.1
Walls, G. Y. (1981). Feeding Ecology of the Tuatara (Sphenodon punctatus) on Stephens Island, Cook Strait. New Zealand Journal of Ecology, 4, 89–97.
Weygoldt, P. (1987). Evolution of parental care in dart poison frogs (Amphibia: Anura: Dendrobatidae). Journal of Zoological Systematics and Evolutionary Research, 25(1), 51–67. https://doi.org/10.1111/j.1439-0469.1987.tb00913.x
Whiting, E. T., & Hastings, A. K. (2014). First Fossil Alligator from the Late Eocene of Nebraska and the Late Paleogene Record of Alligators in the Great Plains. Journal of Herpetology, 49(4), 560–569. https://doi.org/10.1670/14-069
Whiting, S. D., & Whiting, A. U. (2011). Predation by the Saltwater Crocodile (Crocodylus porosus) on Sea Turtle Adults, Eggs, and Hatchlings. Chelonian Conservation and Biology, 10(2), 198–205. https://doi.org/10.2744/CCB-0881.1
Widder, E. A. (1998). A predatory use of counterillumination by the squaloid shark, Isistius brasiliensis. Environmental Biology of Fishes, 53(3), 267–273. https://doi.org/10.1023/A:1007498915860
Wikelski, M., Gall, B., & Trillmich, F. (1993). Ontogenetic changes in food intake and digestion rate of the herbivorous marine iguana (Amblyrhynchus cristatus, Bell). Oecologia, 94(3), 373–379. https://doi.org/10.1007/BF00317112
Wilga, C. D., Motta, P. J., & Sanford, C. P. (2007). Evolution and ecology of feeding in elasmobranchs. Integrative and Comparative Biology, 47(1), 55–69. https://doi.org/10.1093/icb/icm029
Wourms, J. P. (1977). Reproduction and Development in Chondrichthyan Fishes. Integrative and Comparative Biology, 17(2), 379–410. https://doi.org/10.1093/icb/17.2.379
Xavier, F. (1977). An Exceptional Reproductive Strategy in Anura: Nectophrynoides Occidentalis Angel (Bufonidae), an Example of Adaptation to Terrestrial Life by Viviparity. In M. K. Hecht, P. C. Goody, & B. M. Hecht (Eds.), Major Patterns in Vertebrate Evolution (pp. 545–552). Boston, MA: Springer US. https://doi.org/10.1007/978-1-4684-8851-7_19
Xu, X., Zheng, X., & You, H. (2009). A new feather type in a nonavian theropod and the early evolution of feathers. Proceedings of the National Academy of Sciences, 106(3), 832–834. https://doi.org/10.1073/pnas.0810055106
Yokoyama, S., Zhang, H., Radlwimmer, F. B., & Blow, N. S. (1999). Adaptive evolution of color vision of the Comoran coelacanth (Latimeria chalumnae). Proceedings of the National Academy of Sciences of the United States of America, 96(11), 6279–84. https://doi.org/10.1073/pnas.96.11.6279
Zintzen, V., Roberts, C. D., Anderson, M. J., Stewart, A. L., Struthers, C. D., & Harvey, E. S. (2011). Hagfish predatory behaviour and slime defence mechanism. Scientific Reports, 1, 131. https://doi.org/10.1038/srep00131
Zintzen, V., Rogers, K. M., Roberts, C. D., Stewart, A. L., & Anderson, M. J. (2013). Hagfish feeding habits along a depth gradient inferred from stable isotopes. Marine Ecology Progress Series, 485, 223–234. https://doi.org/10.3354/meps10341